




-
中国东部陆架是世界上最平坦宽广的大陆架之一,接受长江等河流巨量的沉积物输入,在东海内陆架形成了长条状分布、厚度达数十米的泥质沉积体[1-2]。前人研究认为东海内陆架泥质区是于冰后期高海面以来逐渐发育而成的,并利用该区快速沉积和沉积体保存良好的优势开展了千年尺度、百年尺度的高分辨沉积记录研究。该地区是研究古海洋、古环境变化的理想地区,获得了丰硕的研究成果[3-4]。该泥质区的形成和发展与长江的物源供给、浙闽沿岸流、台湾暖流和东亚季风密切相关[5-7],同时还接受长江流域内人类活动产生的大量营养盐输入的影响,并最终影响该区域生态环境和生物发育[8-10]。
海洋微体生物对气候变化和海洋环境演变有着敏感的响应。90年代末至今,中国大陆架海域底栖有孔虫与环境变化的研究方兴未艾,取得了丰硕的成果[11-15]。这些研究,主要集中在利用有孔虫组合构建了第四纪以及全新世年代框架、揭示水团演变、分析古环境变化进程[16-18],研究证实有孔虫组合以及有孔虫壳体中的Mg/Ca元素比值、δ 18O稳定同位素组成对水团具有良好的指示作用[19-20]。最近的研究表明,有孔虫组合对长江河口低氧环境亦具有一定的指示作用,可以用于历史时期低氧环境的分析[21-22]。前人研究主要集中在千年尺度有孔虫演化和环境关系方面,而百年来有孔虫演化对于季风、河流输入之间的关系的研究还较为欠缺。本文利用位于东海内陆架泥质区的DH7-2孔岩心沉积物,尝试探讨近两百年以来的浙闽沿海泥质区沉积记录、有孔虫组合变化特征及其对于东亚季风、长江输入的响应问题。
-
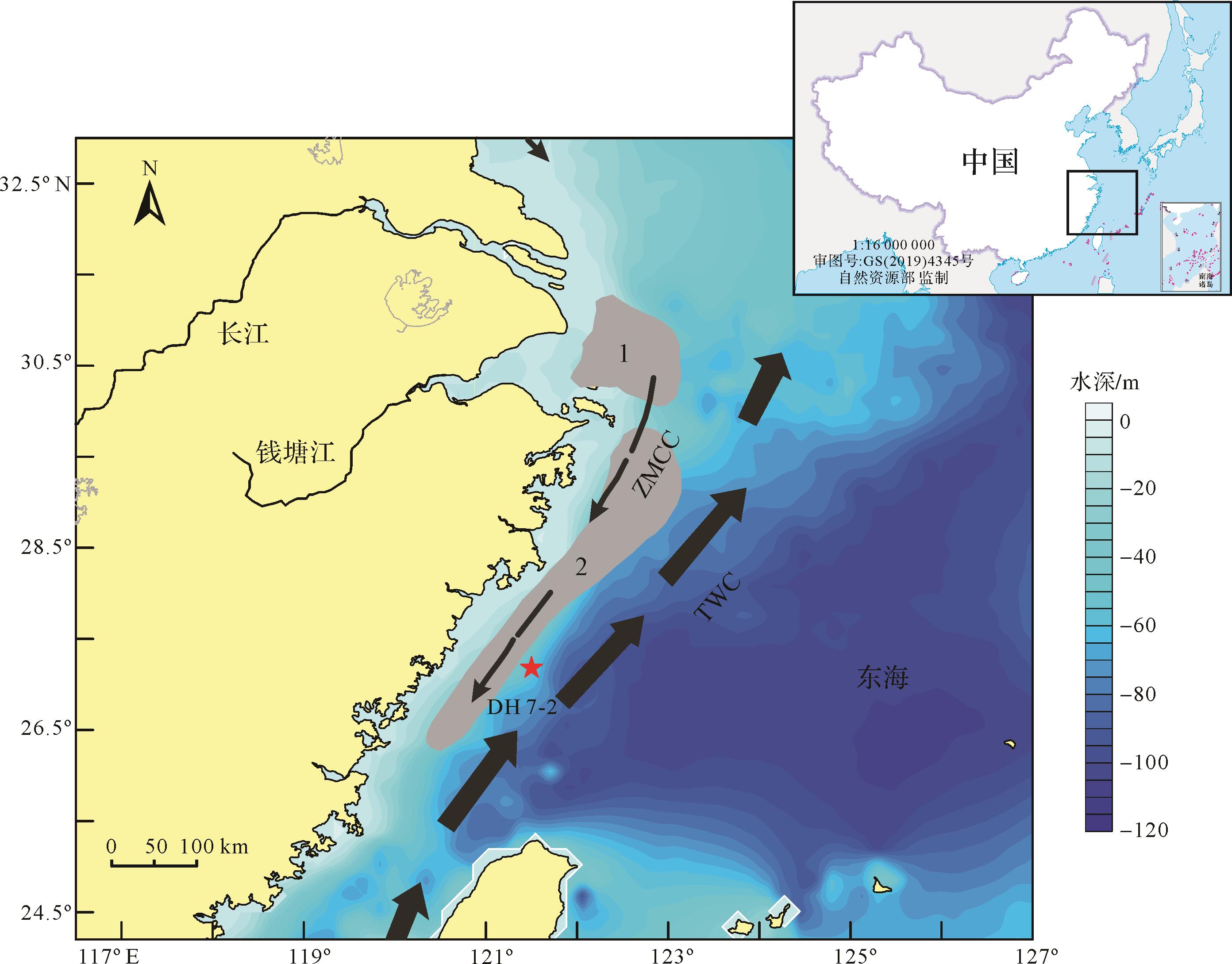
DH7-2岩心由中国海洋大学的“东方红2号”考察船于2011年利用重力取样器采集得到,岩心总长度约193 cm,站位位于东海内陆架泥质区中南部(站位坐标121.50° E,27.18° N),水深58.2 m(图1)。

图 1 DH7⁃2岩心站位及区域环流特征图(改绘自文献[23⁃24];ZMCC:浙闽沿岸流;TWC:台湾暖流;1.长江口泥质区;2.浙闽沿岸泥质区)
Figure 1. Map of the collection site for core DH7⁃2 and local currents (modified from references[23⁃24]; ZMCC: Zhejiang Fujian coastal current; TWC: Taiwan warm current; 1. Changjiang estuary mud area; 2. Zhejiang Fujian coastal mud area)
该岩心为松散沉积物,总体较为均一,以粉砂和黏土为主,黏性强,富含水,上部0~40 cm呈灰黄色,40~68 cm呈灰色略带黄色,68~194 cm呈深灰色,局部可见零星生物碎屑,中部虫孔构造较为发育。在室内对DH7-2岩心进行了室内分样,其中粒度以0.25 cm间隔分样,共分得767个样品;放射性核素测年样品以1 cm间隔进行分样,将分样后的样品装入透明塑料样品袋中密封保存,对其中39个样品进行测试分析;有孔虫样品以5 cm间隔取样,共分析鉴定39个样品。
沉积物粒度测试在中国海洋大学海底科学与探测教育部重点实验室完成,具体步骤是:取大约1 g左右的样品放入50 mL的烧杯中,加入5 mL 30% H2O2以去除有机质,静置24 h,然后加入分散剂(六偏磷酸钠)超声30 min后上机测试。实验仪器为英国Malvern公司生产的Malvern2000型激光粒度仪,仪器测量范围为0.02~2 000 μm,粒级分辨率为0.01 ϕ,重复测量的相对误差<3%;采用矩法计算粒度参数[25]。有孔虫相关实验在中国海洋大学海洋沉积物分析实验室完成,按照微体古生物学研究分析的常规手段进行处理:称取完全烘干后的样品10 g置于500 mL烧杯中,加入50 mL 5%浓度的H2O2浸泡24 h,以便除去样品中的有机质;用0.063 mm的标准铜筛进行冲洗,至样品中的泥质完全去除为止;将筛上的样品使用蒸馏水清洁后转移至50 ℃恒温烘箱中烘干,备用。本文采用的方法是根据汪品先[26]、郑执中等[27]对有孔虫属种的分类方案对DH7-2中39个有孔虫样品进行鉴定及数量统计。
DH7-2岩心的放射性同位素210Pb的测试实验委托中国科学院地理与湖泊研究所的湖泊沉积与环境国家重点实验室完成。首先将约10 g沉积物样品烘干,烘干后装入专用的测试样品盒密封15天,随后对各样品同时进行无损坏的多种核素的直接测量。所用仪器为多道高纯锗井型探测器(OrtecHPGe GWL)γ谱分析系统,过剩210Pb直接由总210Pb扣除226Ra得到。该方法测试相对误差<10%。
-
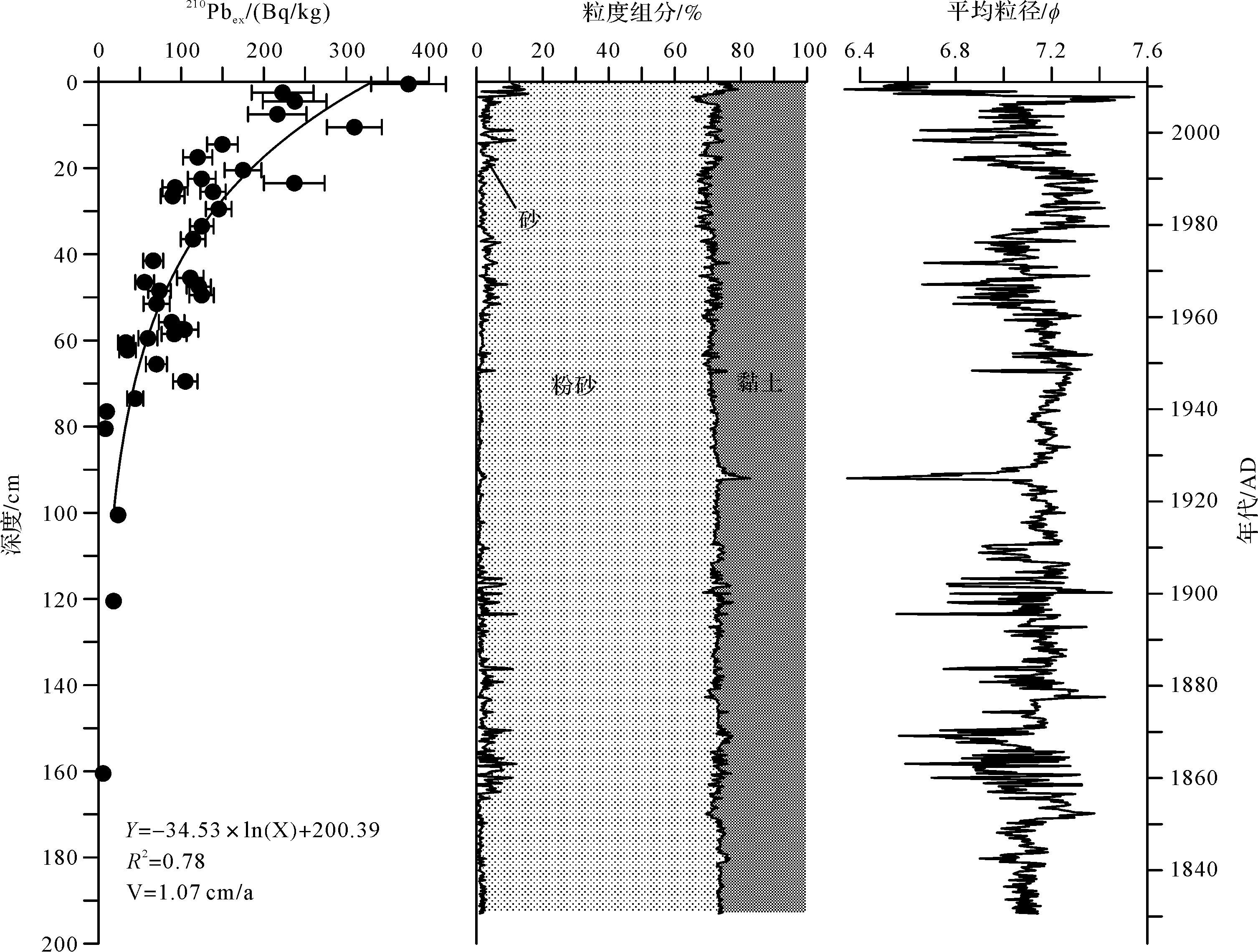
DH7-2岩心210Pbex随深度的分布特征明显(图2),表层无明显扰动现象,在0~80 cm深度区间210Pbex的比活度随深度增加而呈现明显的指数衰减趋势,而80 cm以下层位210Pbex比活度值基本稳定,因而采用0~80 cm的数据进行线性拟合,使用210Pb稳定初始放射性通量(CIC)模式计算平均沉积速率为1.07 cm/a,相关系数R 2为0.78,与邻近海域平均沉积速率较为接近[28]。考虑到内陆架是在冰后期高海平面以来沉积形成的,从高海平面至今该区沉积环境总体上处于较为稳定的状态[29];同时,该岩心的岩性组成变化小,也显示沉积环境较为稳定,所以将210Pb测定的平均沉积速率扩展到整个岩心。

图 2 DH7⁃2岩心210Pbex强度及粒度组成特征
Figure 2. Excess 210Pb profiles with linear fitting and grain size composition of sediment in core DH7⁃2
DH7-2岩心整体从上到下粒度组成变化不大(图2),粉砂平均含量为69.97%,为岩心主要组分,以谢帕德分类命名法命名为黏土质粉砂。DH7-2岩心的平均粒径(Mz)波动范围是6.34~7.54 ϕ,平均值为7.11 ϕ,整体波动不大,仅在92 cm层位及顶部出现突然变粗的现象;分选系数(σ)波动范围是1.31~2.22,平均值为1.65,整体分选差;偏态(Sk)波动范围是-1.66~1.56,平均值为0.77,整体为正偏态;峰态(Sk)波动范围是1.65~2.86,平均值为2.14,峰形较正态分布曲线窄。在垂向变化上,随深度变化出现了两段粒度相对较粗,粒径波动较大的阶段:下部120~170 cm和上部0~70 cm,两段粗颗粒含量明显增加,且平均粒径波动较大,最上层5 cm粗颗粒沉积物明显增加,平均粒径急剧变小;70~120 cm粉砂含量较大,砂含量较小,平均粒径较细,变化较小,在95 cm出现粒度突变层,粉砂含量大,平均粒径较小。
-
本研究对39个沉积物样品中的有孔虫进行了鉴定[26-27],平均每个样品鉴定300枚以上有孔虫,共识别了24 425枚有孔虫壳体, 识别出常见的有孔虫38属61种。统计发现:DH7-2岩心沉积物中,有孔虫绝对丰度在18~2 491枚/g(图3),平均绝对丰度为440枚/g,底栖有孔虫较富集,占有孔虫总数的59~88.6%,平均含量为76.4%,其中玻璃质壳在底栖有孔虫中所占比例最高,在92.1~100%,胶结质次之,为0~7.5%,瓷质壳最低,在0~0.8%。底栖有孔虫的简单分异度(S)在20~40,复合分异度H(S)为2.37~2.82,属种类型变化不大。

图 3 DH7⁃2岩心有孔虫绝对丰度和参数垂向变化特征(S简单分异度;H(S)复合分异度)
Figure 3. Characteristics of foraminifera absolute abundance and parameters in vertical variation of core DH7⁃2
总体上有孔虫壳径较小,碎片较多,相当数量的有孔虫壳体溶蚀作用明显,在岩心下部有孔虫中可见有孔虫壳体内自生黄铁矿填充。这些现象可能是有孔虫死亡后在沉积过程中受到水流搬运发生机械磨蚀出现破损,或者在成岩作用阶段有孔虫受到压实作用、自生矿物交代填充等的影响所致。
DH7-2岩心沉积物中分布较为连续且平均含量超过1%的底栖有孔虫共13种(表1),分别为Bolivina robusta、Bulimina marginata、Hanzawaia nipponica、Ammonia compressiuscula、Ammonia paucilo culata、Ammonia ketienziensis、Elphidium advenum、Hyalinea balthica、Nonionella decora、Textularia spp.、Cavarotalia annectens、Globocassidulina subglobosa、Cibicidoides sp.。其中主要的优势种为B.robusta和B.marginata,其平均含量分别为14.52%和12.17%(图4)。样品中平均丰度较低(<1%),但经常出现的属种有:Nonion belridgensis、Astrononion tasmaniensis、Lagena spp.、Cibicidoides subhaidingerii、Sigmoilopsis asperula、Fissurina lucida、Lenticulina costata等,另外还有一些含量极少的属种,零星分布于个别层位。
表 1 DH7⁃2岩心部分底栖有孔虫平均含量
Table 1.
Average content of benthic foraminifera in core DH7⁃2 有孔虫属种名称 含量/% Bolivina robusta 14.52 Bulimina marginata 12.17 Hanzawaia nipponica 6.76 Ammonia compressiuscula 6.71 Ammonia pauciloculata 6.54 Ammonia ketienziensis 4.3 Elphidium advenum 3.11 Hyalinea balthica 2.87 Nonionella decora 2.58 Textularia spp. 2.11 Cavarotalia annectens 1.89 Globocassidulina subglobosa 1.74 Cibicidoides sp. 1.74 
图 4 DH7⁃2岩心底栖有孔虫相对丰度(%)
Figure 4. Variation of relative abundance of benthic foraminifera (%) with depth in core DH7⁃2
共鉴定出浮游有孔虫5种,分别为Globigerina bulloides、Globigerinoides ruber、Neogloboquadrina dutertrei、Guembelitria vivans和Globorotalia menardii,占全部有孔虫的23.6%。
从有孔虫垂向分布可以看出(图3),有孔虫绝对丰度在70 cm开始迅速增加,在70~40 cm之间丰度较大,在45 cm达到最大值,40~0 cm有孔虫丰度逐渐减小,其中底栖有孔虫相对含量波动较小,仅在25~0 cm含量有所下降,玻璃质壳含量明显减少,而瓷质壳含量相对增加,胶结壳有孔虫在75~35 cm有较明显增加,35~0 cm又相对减少。岩心样品有孔虫的S和H(S)均在120 cm处开始有明显增加,且在120~20 cm之间有孔虫的S和H(S)相对较大,而在190~120 cm之间两者值较小。
-
在DH7-2中发现有14个属种的底栖有孔虫至少在三个样品中的含量大于2%,对其进行R型因子分析(表2),结果显示出3个主要因子,其累计方差贡献为69.25%,基本代表了底栖有孔虫的主要分布。同时,图4中有孔虫在整个岩心中的同步变化分析结果,可以明显地分出3组有相似变化规律的有孔虫,且因子分析结果和组分分析结果具有很好的一致性,因此,把该岩心出现的有孔虫分成3个组合,并与因子分析结果一一对应。
表 2 DH7⁃2底栖有孔虫R型因子载荷矩阵
Table 2.
R⁃type factor load matrix of benthic foraminifera in core DH7⁃2 底栖有孔虫属种名称 成份 因子1(F1) 因子2(F2) 因子3(F3) H.nipponica 0.889 0.098 -0.141 A.compressiuscula 0.884 -0.044 -0.244 Textularia spp. 0.861 0.354 0.116 E.advenum 0.834 -0.251 -0.231 Cibicidoides sp. -0.834 0.054 -0.257 C.subhaidingerii 0.801 0.377 -0.015 G.subglobosa -0.626 0.290 0.291 N.decora -0.609 0.050 -0.330 A.pauciloculata -0.584 0.318 0.058 B.marginata -0.152 -0.899 -0.016 C.annectens -0.115 -0.703 -0.199 H.balthica -0.397 0.678 -0.286 A.ketienziensis -0.031 -0.272 0.843 B.robusta 0.224 0.179 0.598 方差贡献率 40.69% 17.15% 11.43% 组合1:以H.nipponica为主,包括A.compressiu scula、Textularia spp.、E.advenum和C.subhaidingerii等五个属种有孔虫,其中H.nipponica、A.compressiuscula、Textularia spp.和E.advenum在因子1中正载荷较高。组合1中有孔虫相对丰度变化规律均为岩心下部190~120 cm有孔虫相对含量少,120~70 cm相对含量缓慢增加又减少,70~0 cm明显增加。A.compressiuscula、Textularia spp.、E.advenum这些种类的有孔虫均为生活在20~50 m陆架海区的表生属种[30],适应富氧但营养物质较为贫乏的海水环境。
组合2:以B.marginata为主,其次为C.annectens和H.balthica,B.marginata和C.annectens在因子2中表现出较大的负载荷,同时N.decora和Cibicidoides sp.在因子1中也有较大负载荷。组合2在岩心中190~120 cm有孔虫相对丰度较高,并逐渐增加,但是120~70 cm相对含量趋于减少,在70~0 cm含量较少,并保持稳定,该组合中B.marginata是生活在50 m以深的陆架浅海的内生属种,与C.annectens、N.decora 和Cibicidoides sp.三个种一致,在相对水温较暖的低氧富营养盐的环境中较为富集[21,31]。
组合3:以B.robusta和A.ketienziensis为主要有孔虫属种,并且显示了较高的正载荷。该有孔虫组合在整个岩心中都具有较高的相对含量,是该区域的优势属种,在岩心中相对含量的差异不明显,代表了该区域整体的正常海水陆架沉积环境特征。该组合还包括A.pauciloculata,H.balthica和G.subglobosa。B.robusta和A.ketienziensis这两种有孔虫均为内生种属[31],其中,B.robusta是浙闽沿岸泥质区优势种属,多分布在东海陆架,深海和陆架浅海,40~120 m水深较冷水环境,而A.ketienziensis多出现在>50 m水深的浅海环境中。该组中有孔虫都较适应低氧富营养盐的环境。
-
有孔虫丰度变化受到海洋环境变化的强烈影响,而沉积物中有孔虫的变化,可以反映沉积环境的变化[18]。根据有孔虫绝对丰度和底栖有孔虫相对丰度的垂向变化和因子分析结果,可以看出两次较明显的突变,分别位于70 cm和120 cm。岩心样品所含底栖有孔虫组合较为单调,但在70 cm发生了变化,由B.marginata - B.robusta组合变为了B.robusta - H.nipponica组合,从组成变化上看,有孔虫所反映的沉积环境总体呈现缺氧富营养盐状态,但从下向上缺氧状况减缓,营养盐含量相对降低。
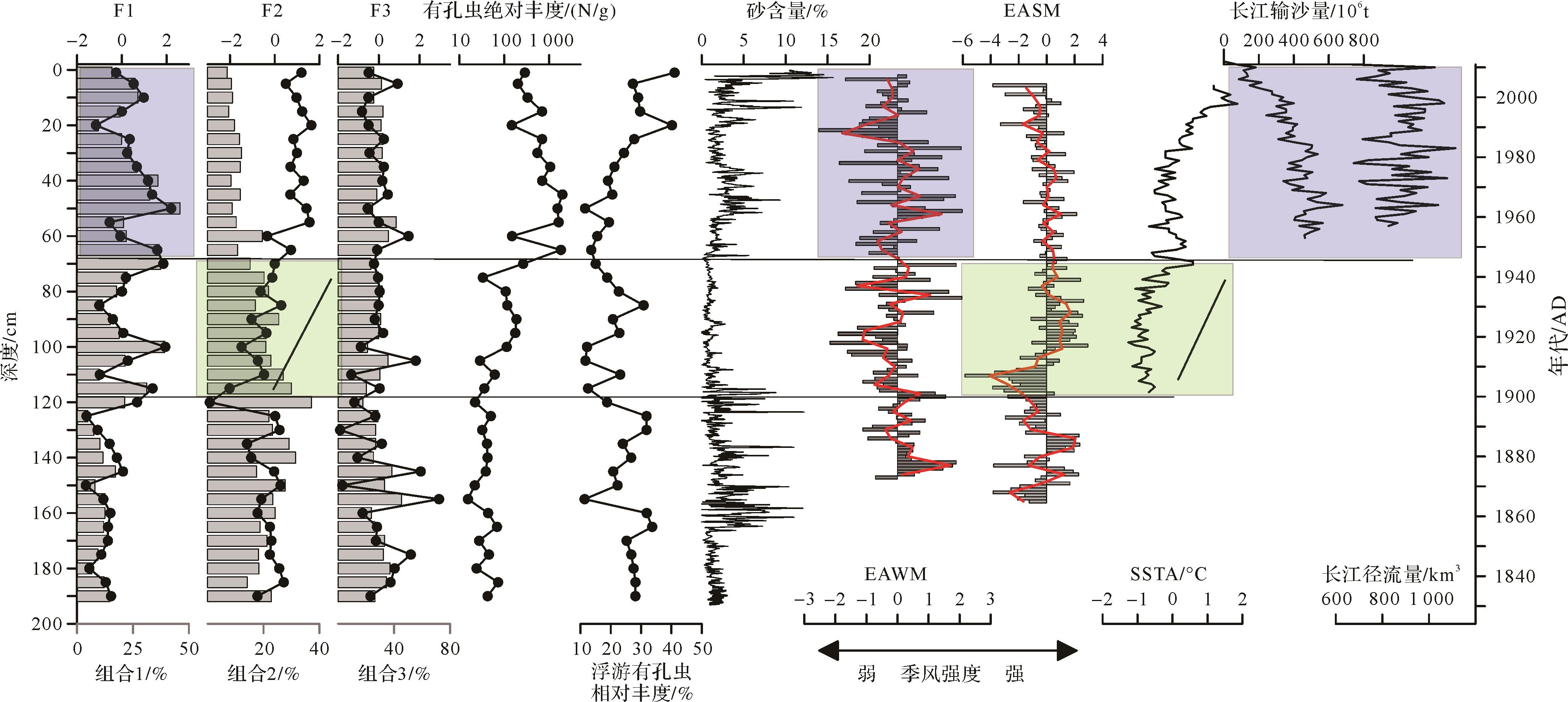
因此,根据以上分析将有孔虫组合变化分为三个阶段(图5)[32-37]:

图 5 DH7⁃2岩心有孔虫组合及主成分变化与沉积环境变化相关指标(EAWM:东亚冬季风[32cm3/g33];EASM:东亚夏季风[34⁃35],红线为平滑曲线;SSTA:东海海水表层温度距平年际变化[36];长江径流量和输沙量数据来自文献[37])
Figure 5. Foraminifera principal component change in core DH7⁃2 and related indices of sedimentary environment change (EAWM: East Asian Winter monsoon[32⁃33]; EASM: East Asian Summer monsoon[34⁃35], red line is the smoothed curve; SSTA: sea surface temperature anomaly[36]; the runoff and sediment discharge data of the Changjiang River are from reference[37])
190~120 cm(1840—1900年):该段内以底栖有孔虫组合1含量最低,组合2、3含量高,因子F1得分低,F2和F3得分较高。其中,适应于富营养盐环境的B.marginata和B.robusta含量最高,平均丰度分别为15.5%和13.7%,这两个种均为内生冷水属种,适应较冷水富氧环境的H.nipponica等有孔虫相对较少。该段内底栖有孔虫绝对丰度整体较低,但有孔虫组合2与组合3相对含量较高,有孔虫适应于富营养盐的环境,并有明显的波动,显示出该段内较为波动的沉积环境。而据气候记载1822—1900年中国处于气候较冷时期,东亚冬季风较强,温度较低,引起沿岸流流速加快,导致沉积物粒度较粗[38],与本研究对比发现DH7-2岩心沉积物粒度在该段内确实存在粒度较粗,且有波动现象,同时,该段浮游有孔虫相对丰度含量较高,由于东海内陆架自生浮游有孔虫可忽略不计,大多数内陆架沉积物中的浮游有孔虫都是潮流从外陆架输入沉积下来,或者外陆架的陆架沉积物再悬浮后被搬运再沉积而来[39]。因此,该段内主要环境影响因素为东亚冬季风较强导致水体混合强烈使得水体含氧量较高[40],同时较强东亚夏季风使得台湾暖流增强[20],携带着营养盐进入东海内陆架,两者共同作用,适应较冷水富氧环境的H.nipponica等有孔虫相对较少,适应于低氧富营养盐环境的B.marginata等有孔虫相对含量较高,但有孔虫绝对丰度较低。
120~70 cm(1900—1945年):该段内底栖有孔虫组合1含量波动较大,相对含量较190~120 cm段增加,组合2有孔虫相对含量由最大逐渐减少,而组合3保持了较高的含量且较稳定,因子F1也有所增加,F2有最低值阶段,同时,底栖有孔虫绝对丰度也呈现增加的趋势。该段仍然以B.robusta和B.marginata含量最高,平均丰度分别为14.8%和14.3%,但是B.marginata相对含量表现为向上明显减少的趋势;组合1中的H.nipponica、A.compressiuscula、Textularia spp.等则出现显著增加趋势(图4)。该阶段组合2有孔虫含量的变化与东亚夏季风强度增加的趋势正好相反,而F2曲线与之吻合,并且与东海SST增加也较吻合。推测该段内在东亚夏季风影响下台湾暖流增强,该区域水温升高,同时带来丰富营养物质,使得底栖有孔虫发育。该段粒度变化较为稳定,以细颗粒沉积为主,表明沉积环境较为稳定,推测是由于东亚冬季风指数较小,变化趋势较为稳定。
70~0 cm(1945—2012年):底栖有孔虫组合1相对含量达到整个岩心的最高值,但向上有孔虫含量逐渐减少,组合2则趋于低值,而组合3变化不大,因子2和因子3相对较高并保持稳定。该段以B.robusta和H.nipponica为主要有孔虫属种,平均丰度为14.6%和9%,对比190~70 cm,底栖有孔虫主要属种发生了明显的改变,由B.marginata-B.robusta组合变为了B.robusta-H.nipponica组合,有孔虫绝对丰度迅速增加,并在该段内有最大丰度,该段平均绝对丰度为964 N/g是190~70 cm整段平均绝对丰度234 N/g的4.1倍,也是胶结质的有孔虫含量较高的部分。该段沉积物粒度平均粒径也变粗,粗粒级组分含量较高,与此同时,长江输沙量逐渐减少(图5),而该阶段东亚冬季风有波动,但是并没有特别的变化,分析认为是长江输沙减少导致的河口和近海沉积物粗化,这也被最近的研究所证实[41];东亚夏季风强度保持稳定,并有逐渐减小的趋势,可能导致台湾暖流逐渐减小,带来的营养盐也逐渐减少。
这个阶段,长江流域范围内人类活动的影响开始显现[42],由长江输入东海的各类营养盐急剧增加,大大促进了有孔虫的发育,且该阶段表层海水温度高,也有利于有孔虫的发育。有孔虫绝对丰度在40 cm处(1970年)达到最大值后又逐渐减小,推测是该时期东亚冬季风和东亚夏季风均由强减弱,冬季风影响下的沿岸流也逐渐减弱,水体混合减弱,含氧量降低,携带物质及营养盐减少,同时东亚夏季风减弱,由台湾暖流带来的营养盐也减少,从而影响有孔虫丰度降低。
对比各指标发现,发生变化层位与粒度组分变化层位较为一致,其中120 cm处的突变,可能与东亚冬季风减弱有关,表现为沉积物粒度由粗变细,水温下降,代表冷水环境的有孔虫明显增加;70 cm处的突变,可能与水温增加[36]和人类活动强烈增加有关[43],有孔虫绝对丰度增加明显,营养开始过剩,低氧程度可能增加,适应生活在较高含氧量的底栖有孔虫属种明显减少。
-
由以上分析可以发现,研究区底栖有孔虫的丰度受到东亚季风强度、河流输入和人类活动等因素的共同影响。DH7-2岩心所在区域受到两侧浙闽沿岸流和台湾暖流的影响,浙闽沿岸流为岩心所在的区域带来丰富的营养盐,而台湾暖流为岩心所在区域带来高温的洋流,通过前人研究可以知道,南下的浙闽沿岸流的输运强度受到东亚冬季风的影响,而台湾暖流主要受到东亚夏季风的影响,因而底栖有孔虫的相对丰度变化在一定程度上受到东亚季风强度演化的综合影响。东亚夏季风对岩心有孔虫的影响主要在190~70 cm段,该段内东亚夏季风与适应暖水环境的有孔虫含量及因子2变化规律较为一致;东亚冬季风较为稳定的变化趋势与代表该区整体沉积环境的有孔虫因子3较为一致。对比之下,东亚季风对底栖有孔虫的影响在1945年后(70~0 cm)显得非常微弱,取而代之的是受人类活动的影响占主导。
历史资料分析表明,自1960年以来,长江口硝酸盐浓度上升10倍,磷酸盐上升2倍,N/P比值从1963年的30,上升至2004年的150,底层水体中溶解氧浓度降低明显[44],而有孔虫绝对丰度自1945年开始迅速增加,认为与长江输入营养盐的剧烈增加有关。自1970年以来,长江口进入富营养盐化状态,赤潮频发,长江口以及邻近海域低氧区逐年扩大[45],河口及近海低氧导致底栖有孔虫受到影响,适宜富氧环境的H.nipponica等属种相对含量自1960年代开始,由最大开始呈下降趋势,说明了人类活动带来的富营养化和水体低氧区域的扩展,而有孔虫绝对丰度也由最大开始减少,同时说明人类活动带来的富营养化和水体低氧化,一定程度地限制了有孔虫的进一步发育。
-
通过东海内陆架沉积物岩心有孔虫组合变化和影响因素的综合分析,认为浙闽沿岸泥质区近200年来有孔虫组合变化可以划分为3个阶段,1822—1900年,有孔虫绝对丰度低,冷水富氧环境的有孔虫较为发育;1900—1945年,有孔虫绝对丰度和适宜暖水环境的有孔虫增加;自1945年以来,有孔虫绝对丰度急剧增加,底栖有孔虫种属组合也出现明显的改变。百年尺度内有孔虫组合的变化主要受区域性气候因素和人类活动双重驱动下该区海水水体含氧量和营养盐状态的制约。1945年之前,该处有孔虫主要受到东亚季风制约,冬季风强劲时水体混合增强,有利于适应富氧环境的底栖有孔虫发育;而夏季风强劲时有利于台湾暖流带来丰富的营养盐,促进暖水富营养盐环境的底栖有孔虫发育。1945年以来,人类活动对有孔虫的影响开始占主导地位,人类活动向河口和近海输入丰富的营养盐大大促进了有孔虫的发育,导致有孔虫丰度峰值的出现,但富营养化和水体低氧区域的扩展则又一定程度地限制了有孔虫的进一步发育。
Foraminiferal Assemblage Changes and Their Influence Factors over the Past 200 Years from the East China Sea
-
摘要:
基于东海内陆架泥质区DH7-2孔岩心,使用激光粒度分析仪、体视显微镜开展了沉积物粒度和有孔虫组成的分析和鉴定,并利用高能γ能谱仪进行了沉积物岩心的210Pb年代标定,探讨了近二百年来有孔虫组合变化及其影响因素。研究表明:近200年来有孔虫组合变化可以划分为3个阶段,1822—1900年,主要受到该时期较强东亚冬季风的影响,有孔虫绝对丰度低,以Bulimina marginata和Bolivina robusta为主,适应于冷水富氧环境的有孔虫较为发育;1900—1945年,主要受到该时期较强东亚夏季风增强的影响,有孔虫绝对丰度和适宜暖水环境的有孔虫增加;自1945年以来,有孔虫绝对丰度急剧增加,种属组合也出现明显的改变,这与该时期大量的营养盐物质输入东海、水体富营养化密切相关。该研究表明1945年之前,东海内陆架海域有孔虫主要受到自然因素的影响,而1945年后受到人类活动的强烈影响。 Abstract:Based on core DH7-2 collected in the inner continental shelf of the East China Sea, we used a laser particle size analyzer to analyze sediment grain size, a microscope to identify foraminifera, and a high-purity γ spectrometer to measure the average deposition rate. Then we discussed the foraminiferal assemblage changes and their influence factors in the past 200 years. The results show that there were three periods in foraminiferal assemblage in the past 200 years. During the period of 1822-1900, the foraminifera were dominated by the relative cold and high oxygen species, mainly including Bulimina marginata and Bolivina robusta, with a low absolute abundance, which was mainly influenced by the East Asian winter monsoon. During the period of 1900-1945, the abundance of foraminifera and the warm species of foraminifera increased, which were mainly affected by the strengthening of the East Asian summer monsoon. Since 1945, the absolute abundance of foraminifera increased dramatically, and the species assemblage changed significantly, which mainly resulted from a large number of nutrients imported into the East China Sea from the Yangtze River related to anthropogenic activities. This study shows that, before 1945, foraminifera in the inner shelf of the East China Sea were mainly controlled by natural processes; meanwhile, human activities play a dominant role in foraminifera developing after 1945. -
图 1 DH7⁃2岩心站位及区域环流特征图(改绘自文献[23⁃24];ZMCC:浙闽沿岸流;TWC:台湾暖流;1.长江口泥质区;2.浙闽沿岸泥质区)
Figure 1. Map of the collection site for core DH7⁃2 and local currents (modified from references[23⁃24]; ZMCC: Zhejiang Fujian coastal current; TWC: Taiwan warm current; 1. Changjiang estuary mud area; 2. Zhejiang Fujian coastal mud area)
图 2 DH7⁃2岩心210Pbex强度及粒度组成特征
Figure 2. Excess 210Pb profiles with linear fitting and grain size composition of sediment in core DH7⁃2
图 3 DH7⁃2岩心有孔虫绝对丰度和参数垂向变化特征(S简单分异度;H(S)复合分异度)
Figure 3. Characteristics of foraminifera absolute abundance and parameters in vertical variation of core DH7⁃2
图 4 DH7⁃2岩心底栖有孔虫相对丰度(%)
Figure 4. Variation of relative abundance of benthic foraminifera (%) with depth in core DH7⁃2
图 5 DH7⁃2岩心有孔虫组合及主成分变化与沉积环境变化相关指标(EAWM:东亚冬季风[32cm3/g33];EASM:东亚夏季风[34⁃35],红线为平滑曲线;SSTA:东海海水表层温度距平年际变化[36];长江径流量和输沙量数据来自文献[37])
Figure 5. Foraminifera principal component change in core DH7⁃2 and related indices of sedimentary environment change (EAWM: East Asian Winter monsoon[32⁃33]; EASM: East Asian Summer monsoon[34⁃35], red line is the smoothed curve; SSTA: sea surface temperature anomaly[36]; the runoff and sediment discharge data of the Changjiang River are from reference[37])
表 1 DH7⁃2岩心部分底栖有孔虫平均含量
Table 1.
Average content of benthic foraminifera in core DH7⁃2 有孔虫属种名称 含量/% Bolivina robusta 14.52 Bulimina marginata 12.17 Hanzawaia nipponica 6.76 Ammonia compressiuscula 6.71 Ammonia pauciloculata 6.54 Ammonia ketienziensis 4.3 Elphidium advenum 3.11 Hyalinea balthica 2.87 Nonionella decora 2.58 Textularia spp. 2.11 Cavarotalia annectens 1.89 Globocassidulina subglobosa 1.74 Cibicidoides sp. 1.74  下载: 导出CSV
下载: 导出CSV
表 2 DH7⁃2底栖有孔虫R型因子载荷矩阵
Table 2.
R⁃type factor load matrix of benthic foraminifera in core DH7⁃2 底栖有孔虫属种名称 成份 因子1(F1) 因子2(F2) 因子3(F3) H.nipponica 0.889 0.098 -0.141 A.compressiuscula 0.884 -0.044 -0.244 Textularia spp. 0.861 0.354 0.116 E.advenum 0.834 -0.251 -0.231 Cibicidoides sp. -0.834 0.054 -0.257 C.subhaidingerii 0.801 0.377 -0.015 G.subglobosa -0.626 0.290 0.291 N.decora -0.609 0.050 -0.330 A.pauciloculata -0.584 0.318 0.058 B.marginata -0.152 -0.899 -0.016 C.annectens -0.115 -0.703 -0.199 H.balthica -0.397 0.678 -0.286 A.ketienziensis -0.031 -0.272 0.843 B.robusta 0.224 0.179 0.598 方差贡献率 40.69% 17.15% 11.43%
下载: 导出CSV
-
[1] 石学法,刘升发,乔淑卿,等. 东海闽浙沿岸泥质区沉积特征与古环境记录[J]. 海洋地质与第四纪地质,2010,30(4):19-30. Shi Xuefa, Liu Shengfa, Qiao Shuqing, et al. Depositional features and palaeoenvironmental records of the mud deposits in MinZhe coastal mud area, East China Sea[J]. Marine Geology & Quaternary Geology, 2010, 30(4): 19-30. [2] 张凯棣,李安春,董江,等. 东海表层沉积物碎屑矿物组合分布特征及其物源环境指示[J]. 沉积学报,2016,34(5):902-911. Zhang Kaidi, Li Anchun, Dong Jiang, et al. Detrital mineral distributions in surface sediments of the East China Sea: Implications for sediment provenance and sedimentary environment[J]. Acta Sedimentologica Sinica, 2016, 34(5): 902-911. [3] Liu S F, Shi X F, Liu Y G, et al. Holocene paleoclimatic reconstruction based on mud deposits on the inner shelf of the East China Sea[J]. Journal of Asian Earth Sciences, 2013, 69: 113-120. [4] Hu B Q, Li J, Zhao J T, et al. Late Holocene elemental and isotopic carbon and nitrogen records from the East China Sea inner shelf: Implications for monsoon and upwelling[J]. Marine Chemistry, 2014, 162: 60-70. [5] 胡敦欣,杨作升. 东海海洋通量关键过程[M]. 北京:海洋出版社,2001:1-204. Hu Dunxin, Yang Zuosheng. The key process of marine fluxes in East China Sea[M]. Beijing: Ocean Press, 2001: 1-204. [6] 刘升发,石学法,刘焱光,等. 近2ka以来东海内陆架泥质区高分辨率的生物硅记录及其古生产力意义[J]. 沉积学报,2011,29(2):321-327. Liu Shengfa, Shi Xuefa, Liu Yanguang, et al. High-resolution record of biogenic silica and its paleoproductivity implication in mud area, East China Sea inner shelf over the last 2000 years BP[J]. Acta Sedimentologica Sinica, 2011, 29(2): 321-327. [7] 李小艳,翦知湣,石学法,等. 东海内陆架泥质沉积记录的全新世快速气候波动[J]. 科学通报,2015,60(4):411-419. Li Xiaoyan, Jian Zhimin, Shi Xuefa, et al. Holocene abrupt climate fluctuations: Sedimentary record from the inner-shelf mud area of the East China Sea[J]. Chinese Science Bulletin, 2015, 60(4): 411-419. [8] Chen L L, Liu J, Xing L, et al. Historical changes in organic matter input to the muddy sediments along the Zhejiang-Fujian Coast, China over the past 160 years[J]. Organic Geochemistry, 2017, 111: 13-25. [9] Chen L L, Liu J, Wang J S, et al. Sources and distribution of tetraether lipids in sediments from the Zhejiang–Fujian coastal mud area, China, over the past 160 years: Implications for paleoclimate change[J]. Organic Geochemistry, 2018, 121: 114-125. [10] Yao P, Zhao B, Bianchi T S, et al. Remineralization of sedimentary organic carbon in mud deposits of the Changjiang Estuary and adjacent shelf: Implications for carbon preservation and authigenic mineral formation[J]. Continental Shelf Research, 2014, 91: 1-11. [11] 徐勇航,陈坚,王爱军,等. 南海北部陆架中全新世沉积记录及古环境意义[J]. 沉积学报,2012,30(3):556-564. Xu Yonghang, Chen Jian, Wang Aijun, et al. Records and plaeoenvironmental implications from the shelf of the northern South China Sea since the Mid-Holocene[J]. Acta Sedimentologica Sinica, 2012, 30(3): 556-564. [12] 史光辉. 东海陆架泥质区底栖有孔虫记录及其环境意义[D]. 青岛:中国海洋大学,2013. Shi Guanghui. Benthic foraminifera response to changes in paleoenviornments of mud area on the East China Sea shelf[D]. Qingdao: Ocean University of China, 2013. [13] Lei Y, Li T G. Atlas of benthic foraminifera from China Seas[M]. Berlin Heidelberg: Springer, 2016: 1-399. [14] Du S H, Li B S, Chen M H, et al. Paleotempestology evidence recorded by eolian deposition in the Bohai Sea coastal zone during the last interglacial period[J]. Marine Geology, 2016, 379: 78-83. [15] 陈晓辉,李日辉,蓝先洪,等. MIS3中期以来北黄海中部陆架古环境演化[J]. 沉积学报,2016,34(1):102-110. Chen Xiaohui, Li Rihui, Lan Xianhong, et al. Paleo-environmental evolution in the central shelf of the North Yellow Sea since mid-MIS3[J]. Acta Sedimentologica Sinica, 2016, 34(1): 102-110. [16] 庄丽华,李铁刚,常凤鸣,等. 东海中陆架晚第四纪底栖有孔虫定量分析[J]. 海洋地质与第四纪地质,2004,24(1):43-50. Zhuang Lihua, Li Tiegang, Chang Fengming, et al. Quantitative analysis on the benthic foraminifera fauna in Late Quaternary period in the north continental shelf of the East China Sea[J]. Marine Geology & Quaternary Geology, 2004, 24(1): 43-50. [17] 赵泉鸿,翦知湣,张在秀,等. 东海陆架泥质沉积区全新世有孔虫和介形虫及其古环境应用[J]. 微体古生物学报,2009,26(2):117-128. Zhao Quanhong, Jian Zhimin, Zhang Zaixiu, et al. Holocene benthic foraminifera and ostracoda from the shelf mud area of the East China Sea and their paleoenvironmental implications[J]. Acta Micropalaeontologica Sinica, 2009, 26(2): 117-128. [18] 李小艳,翦知湣,石学法,等. 全新世东海内陆架泥质区有孔虫特征及其古环境意义[J]. 海洋地质与第四纪地质,2012,32(4):61-71. Li Xiaoyan, Jian Zhimin, Shi Xuefa, et al. Holocene foraminifera from the mud area of the inner shelf, East China Sea and their paleoenvironmental significance[J]. Marine Geology & Quaternary Geology, 2012, 32(4): 61-71. [19] Xiang R, Yang Z S, Saito Y, et al. Paleoenvironmental changes during the last 8400 years in the southern Yellow Sea: Benthic foraminiferal and stable isotopic evidence[J]. Marine Micropaleontology, 2008, 67(1/2): 104-119. [20] 张在秀,赵泉鸿,陈荣华,等. 东海泥质沉积区8千年来的有孔虫Mg/Ca和δ18O记录及其古环境意义[J]. 海洋地质与第四纪地质,2010,30(3):79-86. Zhang Zaixiu, Zhao Quanhong, Chen Ronghua, et al. Foraminiferal Mg/Ca and δ18O records in the mud aera of the East China Sea during the last 8 000 years and their palaeoenvironmental implications[J]. Marine Geology & Quaternary Geology, 2010, 30(3): 79-86. [21] 蔡庆芳,王飞飞,印萍,等. 全新世以来长江口底栖有孔虫组合及古环境演化[J]. 海洋地质与第四纪地质,2016,36(6):175-184. Cai Qingfang, Wang Feifei, Yin Ping, et al. Subassemblages of benthic foraminifera and palaeoenvironmental implication in the Yangtze River Estuary since Holocene[J]. Marine Geology & Quaternary Geology, 2016, 36(6): 175-184. [22] 蔡庆芳,王飞飞,印萍,等. 长江口低氧区底栖有孔虫组合[J]. 海洋地质前沿,2013,29(6):44-51. Cai Qingfang, Wang Feifei, Yin Ping, et al. Benthic foraminiferal assemblages in the hypoxic area of Yangtze River Estuary[J]. Marine Geology Frontiers, 2013, 29(6): 44-51. [23] 李广雪,杨子赓,刘勇. 中国东部海域海底沉积环境成因研究[M]. 北京:科学出版社,2005:1-65. Li Guangxue, Yang Zigeng, Liu Yong. Origins of seafloor sedimentary environments in Eastern China Sea regions[M]. Beijing: Science Press, 2005: 1-65. [24] Wang F F, Yu Z G, Xu B C, et al. Nepartak typhoon influenced bottom sediments from the Yangtze River Estuary and Adjacent East China Sea-foraminiferal evidence[J]. Geochemistry, Geophysics, Geosystems, 2018, 19(4): 1049-1063. [25] McManus J. Grain size determination and interpretation[M]//Tucker M E. Techniques in sedimentology. Oxford: Blackwell, 1988. [26] 汪品先. 东海底质中的有孔虫和介形虫[M]. 北京:海洋出版社,1988:1-438. Wang Pinxian. Foraminifera and ostracoda in bottom sediments of the East China Sea[M]. Beijing: Ocean Press, 1988: 1-438. [27] 郑执中,郑守仪. 黃海和东海的浮游有孔虫[J] . 海洋与湖沼,19603,(3):125-152,218-229. Cheng Tsichung, Cheng Sauyee. The planktonic foraminifera of the Yellow Sea and the East China Sea[J]. Oceanologia et Limnologia Sinica, 19603,(3): 125-152,218-229. [28] Qiao S Q, Shi X F, Wang G Q, et al. Sediment accumulation and budget in the Bohai Sea, Yellow Sea and East China Sea[J]. Marine Geology, 2017, 390: 270-281. [29] 王可,郑洪波,Prins M,等. 东海内陆架泥质沉积反映的古环境演化[J]. 海洋地质与第四纪地质,2008,28(4):1-10. Wang Ke, Zheng Hongbo, Prins M, et al. High-resolution paleoenvironmental record of the mud sediments of the East China Sea inner shelf[J]. Marine Geology & Quaternary Geology, 2008, 28(4): 1-10. [30] 李日辉,孙荣涛,徐兆凯,等. 黄海与渤海交界区附近表层沉积物中的底栖有孔虫分布与环境因素制约[J]. 海洋地质与第四纪地质,2014,34(3):93-103. Li Rihui, Sun Rongtao, Xu Zhaokai, et al. Distribution of benthic foraminifera in surface sediments in the junction of Bohai Sea and Yellow Sea and environmental constraints[J]. Marine Geology & Quaternary Geology, 2014, 34(3): 93-103. [31] 程广芬. 厌氧沉积中有孔虫组合特征[J]. 山东海洋学院学报,1988,18(1):70-80. Cheng Guangfen. The characteristics of foraminiferal assemblage in the anoxic sediments[J]. Journal of Shandong College of Oceanology, 1988, 18(1): 70-80. [32] Qiao S Q, Yang Z S, Liu J P, et al. Records of Late-Holocene East Asian winter monsoon in the East China Sea: Key grain-size component of quartz versus bulk sediments[J]. Quaternary International, 2011, 230(1/2): 106-114. [33] Zeng G, Wang W C, Sun Z B, et al. Atmospheric circulation cells associated with anomalous East Asian winter monsoon[J]. Advances in Atmospheric Sciences, 2011, 28(4): 913-926. [34] 施能,鲁建军,朱乾根. 东亚冬、夏季风百年强度指数及其气候变化[J]. 南京气象学院学报,1996,19(2):168-177. Shi Neng, Lu Jianjun, Zhu Qiangen. East Asian winter/summer monsoon intensity indices with their climatic change in 1873~1989[J]. Journal of Nanjing Institute of Meteorology, 1996, 19(2): 168-177. [35] Wang H J, Yang Z S, Wang Y, et al. Reconstruction of sediment flux from the Changjiang (Yangtze River) to the sea since the 1860s[J]. Journal of Hydrology, 2008, 349(3/4): 318-332. [36] 张秀芝,裘越芳,吴迅英. 近百年中国近海海温变化[J]. 气候与环境研究,2005,10(4):799-807. Zhang Xiuzhi, Qiu Yuefang, Wu Xunying. The long-term change for sea surface temperature in the last 100 years in the offshore sea of China[J]. Climatic and Environmental Research, 2005, 10(4): 799-807. [37] 水利部长江水利委员会. 长江泥沙公报2018[R]. 武汉:长江出版社,2019:1-51. Changjiang Water Resources Commission of the Ministry of Water Resources. Changjiang sediment bulletin 2018[R]. Wuhan: Changjiang Press, 2019: 1-51. [38] 程芳晋,俞志明,宋秀贤. 东海近岸泥质区柱状沉积物的百年内沉积粒度变化及其影响因素[J]. 海洋科学,2013,37(10):58-64. Cheng Fangjin, Yu Zhiming, Song Xiuxian. Variations of sediment grain size of the coastal mud area of the East China Sea and the influence factors during recent hundred years[J]. Marine Sciences, 2013, 37(10): 58-64. [39] Yamasaki M, Oda M. Sedimentation of planktonic foraminifera in the East China Sea: Evidence from a sediment trap experiment[J]. Marine Micropaleontology, 2003, 49(1/2): 3-20. [40] 皮仲,李铁刚,南青云. 中全新世以来南黄海岩心记录的沉积环境演变对东亚季风的响应[J]. 海洋地质前沿,2016,32(7):1-10. Pi Zhong, Li Tiegang, Qingyun Nan. Environmental changes since Mid Holocene revealed by core Z1 in the South Yellow Sea and their response to East Asian Monsoon[J]. Marine Geology Frontiers, 2016, 32(7): 1-10. [41] Yang H F, Yang S L, Meng Y, et al. Recent coarsening of sediments on the southern Yangtze subaqueous delta front: A response to river damming[J]. Continental Shelf Research, 2018, 155: 45-51. [42] Sun X S, Fan D J, Liu M, et al. Persistent impact of human activities on trace metals in the Yangtze River Estuary and the East China Sea: Evidence from sedimentary records of the last 60 years[J]. Science of the Total Environment, 2019, 654: 878-889. [43] 刘莹,翟世奎,李军. 长江口与闽浙沿岸泥质区现代沉积记录及其影响因素[J]. 海洋地质与第四纪地质,2010,30(5):1-10. Liu Ying, Zhai Shikui, Li Jun. Depositional records in the mud areas of Changjiang Estuary and off Min-Zhe coast and their influence factors[J]. Marine Geology & Quaternary Geology, 2010, 30(5): 1-10. [44] 柴超. 长江口水域富营养化现状与特征研究[D]. 青岛:中国科学院研究生院(海洋研究所),2006. Chai Chao. Study on the status and characteristics of eutrophication in the Yangtze River Estuary[D]. Qingdao: Institute of Oceanology, Chinese Academy of Sciences, 2006. [45] 吴在兴,俞志明,宋秀贤,等. 基于水质状态和生态响应的综合富营养化评价模型:以山东半岛典型海域富营养化评价为例[J]. 海洋与湖沼,2014,45(1):20-31. Wu Zaixing, Yu Zhiming, Song Xiuxian, et al. A new system of eutrophication assessment for both water quality and ecological response: A case study in typical areas off Shandong peninsula[J]. Oceanologia et Limnologia Sinica, 2014, 45(1): 20-31. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 647
- HTML全文浏览量: 177
- PDF下载量: 75
- 被引次数: 0

