



-
温度是地球表层环境的重要参数之一,重建地质历史时期海水温度对于理解环境特征及其演化具有重要的科学价值[1]。目前,多种地球化学指标或参数已被用于地质历史时期海水温度重建,例如生物壳体δ 18O[2-4]、碳酸盐团簇同位素∆47 [5-7]、有孔虫Mg/Ca比值[8]、C86四醚指数TEX86 [9-10]、长链烯酮不饱和度
牙形刺在寒武系至三叠系海相地层里分布广泛、种类繁多、特征显著、演化迅速,是古生代地层划分与对比的重要标准化石[26-29]。牙形刺主要由磷灰石(即磷酸钙)有序排列而成,结构致密,能够较好抵抗后期成岩作用改造,化学稳定性好[30-31]。重要的是,磷酸盐氧同位素对环境水体温度变化比较敏感[15,32-33],且易于测量,具备开发为古温度计的必要条件。21世纪以来,随着地球化学分析测试方法与技术的快速发展,研究人员持续开发、改进和优化牙形刺氧同位素古温度计[31,34-37],并取得重要进展和认识。例如,通过激光剥蚀电感耦合等离子体质谱仪(LA-ICP-MS)研究揭示,牙形刺不同组织部位抵抗成岩作用改造的能力各异[38];通过透射电镜研究揭示,单颗粒牙形刺内部不同组织部位在晶体大小、形状和晶间(内)孔隙等微观结构上具有差异性[39];通过二次离子质谱仪(SIMS)对牙形刺进行原位微区氧同位素分析,可显著提升古海水温度重建的质量和时间分辨率[35,40]。目前,牙形刺氧同位素已逐渐成为重建深时海水温度的重要手段,得到快速发展和广泛应用,并极大促进了我们对深时地球环境与生物演化的认识。
在此背景下,本文对牙形刺基本特征、磷酸盐氧同位素古温度计原理、分析测试方法与技术,以及古海水温度定量重建影响因素等方面,进行了简要回顾和介绍。同时,进一步讨论了牙形刺氧同位素古温度计的应用现状,并展望其未来发展,以期推动相关研究。
-
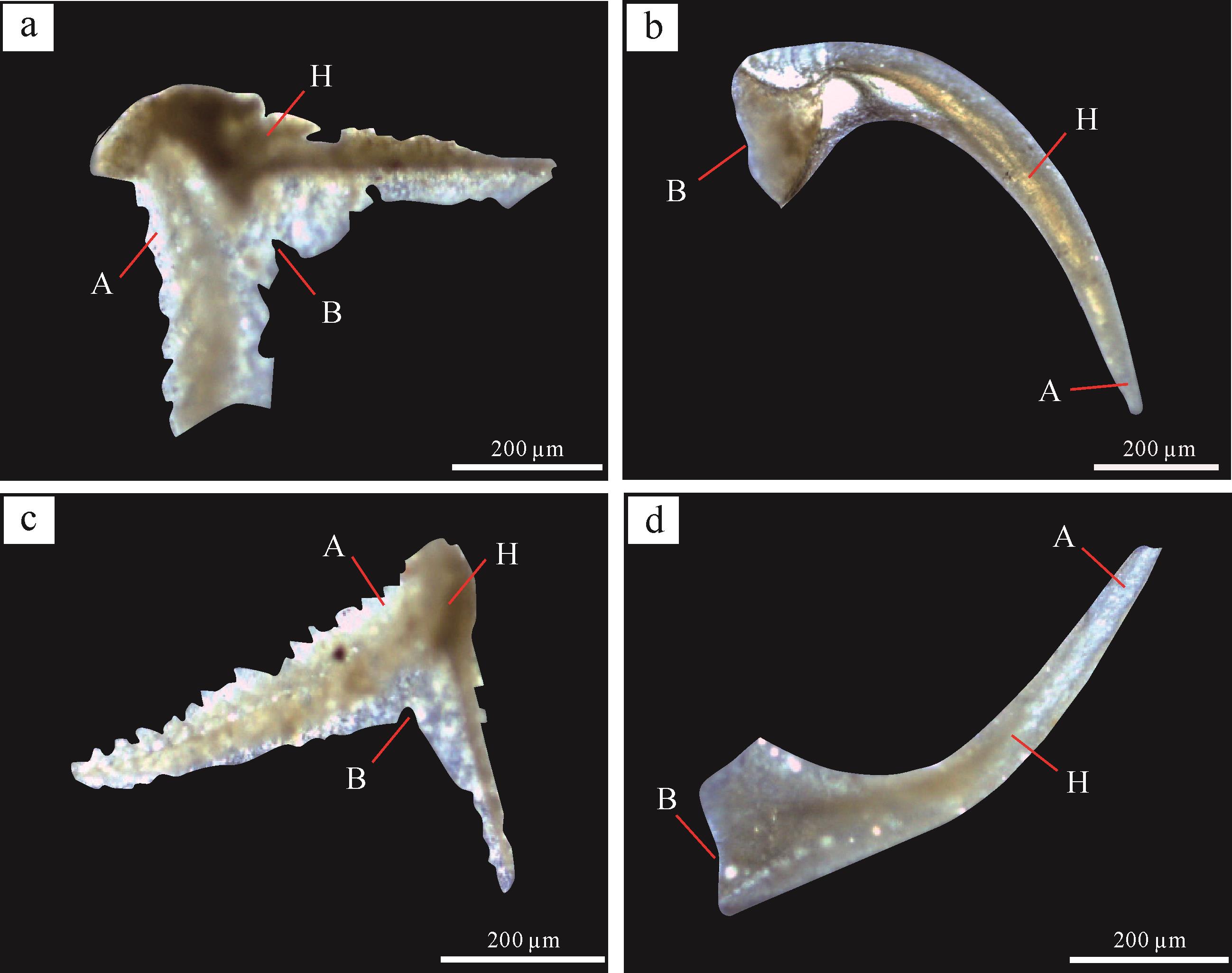
1856年,俄国学者Pander在下奥陶统含海绿石砂岩里首次发现了多种呈单锥状或锥齿状的微体生物结构,并创立了“Conodont”一词[41],即牙形刺(或称牙形石)。牙形刺个体微小,长约0.1~3 mm(表1)。成分上,牙形刺主要成分为细晶碳氟磷灰石,可表达为化学式Ca5Na0.14(PO4)3.01(CO3)0.16F0.73(H2O)0.85 [42]。形貌上,牙形刺通常呈琥珀色、浅黄色或灰黑色,并可细分为单锥、复合和平台型三种形态类型。结构上,牙形刺可分为基腔(basal body)和齿冠(主齿)(crown)两部分[43],其中齿冠可再细分为半透明的玻璃质(hyaline)和不透明的瓷白质(albid)两部分(图1)。生物学上,牙形刺可能属于底栖或分层游泳无颌类脊椎动物的滤食器官部位[44-48],在浅海陆架至深水盆地环境广泛分布,并常见于碳酸盐岩地层。据统计,牙形刺最早出现于早寒武世,并在早—中奥陶世(约140属种)和中泥盆世(约160属种)分别经历了两次辐射高峰,自早石炭世开始逐渐衰减,至晚古生代冰期发生骤减(少于40属种),最终于三叠纪末期绝灭[44,49]。
表 1 牙形刺的基本特征
Table 1.
The main characteristics of conodonts 成分 Ca5Na0.14(PO4)3.01(CO3)0.16F0.73(H2O)0.85 矿物 碳氟磷灰石 颜色 透明琥珀色、浅黄色或灰黑色等 形貌类型 单锥型、复合型和平台型 结构特征 基腔、齿冠(主齿) 寄主生物 底栖或分层游泳动物 时代分布 早寒武世—晚三叠世 生活环境 浅海陆架—深水盆地 
图 1 牙形刺的形态和结构特征
Figure 1. Morphological and structural features of conodonts
牙形刺中氧的赋存状态及稳定性,对于氧同位素古温度计开发和应用至关重要。理论上,牙形刺中约90%~95%的氧主要赋存于PO4 3-[50],其余少量赋存于CO
-
由于生物矿化成因的磷酸盐矿物具有结构和化学稳定性的优势,研究人员不断探索和开发其氧同位素古温度计,使之日益成熟[32-33,37,56]。理论上,水体溶解态磷酸盐氧同位素组成由水体温度及水分子氧同位素值共同决定[15]:温度越高,溶解态磷酸盐和水分子间的氧同位素分馏系数越小,氧同位素组成越偏轻[57]。生物利用海水磷酸盐矿化生成磷灰石,因此可记录同时期海水温度。需要强调的是,在生物矿化过程中,牙形刺氧同位素组成与同时期海水中水分子在何种程度达到热力学平衡分馏,是否存在同牙形动物生命活动相关的动力学分馏(即生命效应,vital effects),仍无定论[25]。目前,牙形刺作为已绝灭生物的磷酸盐质器官,通常通过借鉴基于现生生物所标定的磷酸盐氧同位素—温度经验公式,定量重建古海水温度。研究人员基于特定生物及其生存的一系列水体温度条件(T),测量发现生物矿化成因磷酸盐矿物(δ 18Ophosphate)及周围环境水体(δ 18Ow)的氧同位素组成通常呈现较好的线性关系,因此采用数学方法进行线性拟合,即可获得温度T关于Δ(Δ=δ 18Ophosphate-δ 18Ow)的经验公式。在此背景下,Longinelli et al.[56]针对含磷酸盐的藤壶和软体动物壳,Kolodny et al.[32]针对磷酸盐质鱼齿和鱼骨,Pucéat et al.[33]针对人工饲养的海鲤牙齿等,先后开展了一系列研究,采用不同的样品处理手段和磷酸盐标样参考值,依次建立了多种磷酸盐氧同位素—温度经验公式(表2),并被用于牙形刺氧同位素古温度定量重建。
表 2 常用的磷酸盐氧同位素—温度经验公式
Table 2.
Empirical formulas for paleotemperature reconstruction based on phosphate oxygen isotopes 温度计算经验公式 温度范围/℃ 研究对象 参考文献 T(℃)=111.4-4.3(δ 18Ophosphate-δ 18Ow) 3.5~27.3 藤壶和软体动物壳中的磷酸盐 Longinelli et al.[56]、Grossman[15] T(℃)=113.3-4.38(δ 18Ophosphate-δ 18Ow) 3.5~25 磷酸盐质鱼齿和鱼骨 Kolodny et al.[32]、Grossman[15] T(℃)=118.7-4.22[δ 18Ophosphate+(22.6-δ 18ONBS120c)-δ 18Ow] 3.5~28 磷酸盐质鱼齿 Pucéat et al.[33]、Grossman[15] T(℃)=117.4(±9.5)-4.50(±0.43)(δ 18Ophosphate-δ 18Ow) — 舌形贝和鲨鱼牙齿中的磷酸盐 Lécuyer et al.[37] -
牙形刺氧同位素分析测试方法经过多年的探索和开发,不断朝着低样品量、快速高效、高精准度、高分辨率及原位微区的测试方向发展。牙形刺通常以微小个体弥散分布于沉积岩中,丰度很低,通常需要通过物理和化学手段对其进行分选和提取,以便进行氧同位素等各类分析测试。对于碳酸盐岩样品,通常采用醋酸溶解法,依次进行碎样、酸溶、过筛、重液分离和镜下挑选等流程,实现对牙形刺的分离与收集(图2)。对于页岩样品,碎样后,通常采用直接水煮方式将页岩分解成泥浆,再进行过筛将牙形刺与岩石基质分离;然后,继续进行重液分离和镜下挑选。最后,通过立体显微镜对牙形刺进行属种鉴定和拍照,并选取保存较完好的特定属种进行氧同位素分析。

图 2 基于TC⁃EA⁃IRMS和SIMS方法的牙形刺氧同位素分析测试流程示意图
Figure 2. Analytical workflow diagram of conodont oxygen isotopes through thermal conversion⁃elemental analyzer⁃isotope ratio mass spectrometry (TC⁃EA⁃IRMS) and secondary ion mass spectrometry (SIMS) methods
牙形刺氧同位素古温度计核心在于准确测量其矿物晶格中PO
近年,热转换元素分析—气体同位素质谱法(TC-EA-IRMS方法)逐渐成为牙形刺氧同位素测试的常用方法。该方法首先将牙形刺进行酸溶,释放PO
目前诸多研究发现,牙形刺不同属种或单颗粒不同部位均可存在一定程度的组构和组分差异,导致具有非均一的氧同位素组成特征[36,38]。因此,针对特定属种牙形刺的特定部位,高效而精确提取磷酸盐组分原生氧同位素信号,是牙形刺氧同位素古温度计应用的关键技术。随着原位微区分析测试技术与方法的进步,LA-ICP-MS[76]、SIMS[35,40,77-79]等原位微区分析技术已不断应用于牙形刺氧同位素分析。LA-ICP-MS方法由于激光束斑直径(超过100 μm)较大、空间分辨率较低,难以在单颗牙形刺内部多次采点分析,制约了分析测试数据的精度[76]。相比之下,SIMS方法的离子束斑较小(10 μm)、空间分辨率高,可对单颗牙形刺的不同部位进行高灵敏度的氧同位素分析,同时具有测试快捷、样品需求量小(1~3 ng)、检出限低等优势,逐渐成为分析测试的优选手段。SIMS方法首先筛选特定属种牙形刺进行制靶,然后利用Cs+一次离子束轰击样品表面,生成的二次离子进入质谱仪分析18O/16O比值[80-81],氧同位素测试精度可达0.20‰~0.28‰(1σ)[35,82]。理论上,SIMS分析方法无法有效区分牙形刺中各类含氧组分(PO
-
牙形刺氧同位素古温度计基于生物矿化成因磷酸盐组分与同时期海水氧同位素组成,通过氧同位素—温度经验公式(表2),定量重建古海水温度[32-33,56]。事实上,除了分析测试方法相关的误差之外,牙形动物的生物习性、牙形刺成岩作用、数值量化计算等因素,均可能不同程度地影响该古温度计的精准度及其地质意义。
-
牙形刺氧同位素古海水温度重建,必须考虑牙形动物的生物相或者生物习性[84-85]。牙形动物的生物相是指具有一定沉积环境和生态特征的牙形刺组合,常依据丰度最高的牙形刺属种名称进行命名[86]。例如,Rasmussen et al.[87]基于多元统计学分析,将波罗的陆块中奥陶世牙形刺生物相按照由浅至深的水深顺序依次划分为Baltoniodus-Microzarkodina生物相(内陆架)、Periodon生物相(陆架边缘)和Protopanderodus生物相(远端陆架)。其中,陆架边缘Periodon生物相划分的合理性可得到牙形刺氧同位素证据的支持[85]。由于海水温度存在季节、水深和纬度差异,牙形动物不同属种或底栖[88]、或浮游生活在不同纬度、自表层至数百米不同水深的海洋环境[60,89-90],其生物习性差异导致可能记录不同海水环境的温度信息[91]。因此,同一时期(层位)、不同属种牙形刺的氧同位素组成是否存在属种间差异,会影响古海水温度的重建结果。例如,Buggisch et al.[92]通过对奥陶纪牙形刺研究发现,不同水深环境不同属种牙形刺的氧同位素组成不存在显著差别。此外,Joachimski et al.[60]通过系统比较Frasnian-Famennian界线附近Polygnathus、Icriodus和Palmatolepis三个属的牙形刺氧同位素组成,也未发现明显的差别;然而华南地区同一层段的研究发现,Palmatolepis属牙形刺的δ 18O值要比前两者分别偏低0.6‰~0.7‰和0.5‰~0.9‰[90]。事实上,诸多研究发现,同一时期(层位)牙形刺不同属种之间的δ 18O差值可达约0.5‰~1.1‰[93-94]。这种差异可能源于:1)不同属种牙形动物记录了不同水深环境的温度信号;2)牙形刺氧同位素或存在属种间的生命效应。另一方面,牙形刺氧同位素可能还存在“属种内差异”。据报道,同一地区奥陶系相同层位同一属种、不同个体的牙形刺瓷白质齿冠的δ 18O平均值可相差约1.5‰(VSMOW),体现了部分牙形动物属种的生物习性复杂性[85]。此外,近年有研究发现,基于牙形刺氧同位素所推测的水深与其赋存围岩的沉积水深有时并不相符[58,85,90],二者关系仍需进一步探讨。因此,明确牙形动物的生物相或生物习性,尽量选取特定单一、浮游相属种牙形刺进行氧同位素分析,有助于更精准解译古海水温度变化趋势、更合理解读地质意义。未来,进一步厘定和完善各时代牙形动物的生态分布模型,有助于针对性地挑选适宜的牙形刺属种,优化重建古海水温度曲线。
-
牙形刺氧同位素古海水温度重建,必须考虑成岩作用对其组构和组分的改造影响。传统研究认为,牙形刺个体的玻璃质和瓷白质齿冠能较好的保存初始氧同位素信号[35,73]。然而,牙形刺虽然结构致密,但是在一定的成岩作用下,其初始氧同位素组成仍可能会发生改变[95]。因此,评估牙形刺样品的保存情况和成岩作用改造程度,是进行氧同位素分析测试和古温度精确重建的必要前提。
目前,色变指数(Color Alteration Index,CAI)被广泛用于评估牙形刺所经历的最高埋藏温度及成岩作用改造程度。该指数基于实验条件,按照牙形刺遭受热成岩作用改造强度由弱到强的顺序,依次将其对应颜色量化划分为1~8等级[96-97]:成岩作用温度越高,则牙形刺CAI值越大。通过对比蒙赛尔颜色图(Munsell color chart),可以定性判别牙形刺CAI值[98]。有研究认为,牙形刺CAI值不超过5,其氧同位素组成通常保存较好[18,21,60]。该指标是评估牙形刺遭受成岩作用改造程度的常用指标,然而其依赖于主观视觉判断,且难以客观精确量化。另一方面,Shemesh[99]提出以磷灰石结晶度作为判断其遭受成岩作用改造程度的指标:结晶度越低,则改造程度越弱。随后,Pucéat et al.[100]基于拉曼光谱研究进一步建立了磷灰石结晶度定量化指标(crystallinity index,CI);然而,该指标与初始海水信号保存程度的对应关系较复杂,难以准确指示样品化学组分遭受成岩作用改造的程度。基于扫描电子显微镜(SEM)观察牙形刺磷灰石的形貌和表面结晶特征,可以直观定性评估其遭受重结晶和后期改造的程度[90,95]。此外,部分研究者尝试通过稀土配分模式(REE)和阴极发光(CL)特征来判断牙形刺的保存情况,但是同样难以有效评估其初始氧同位素信号的保存程度[101]。最新研究表明,牙形刺中有机质随着热成熟度不断增加而逐渐向石墨转变,据此可通过拉曼光谱定量评估牙形刺中有机质热演化程度,进而评估其遭受成岩作用改造的程度[102]。基于拉曼光谱评估方法针对加拿大密西西比阶—上三叠统牙形刺样品所估算的最高埋藏温度范围,与CAI指标的估算结果总体比较相近[103],支持了该方法的有效性。实践中,牙形刺瓷白质齿冠被认为在成岩改造过程中具有更好的结构和化学稳定性,常作为氧同位素信号提取的首选部位。然而也有研究报道,牙形刺瓷白质齿冠部位相对于玻璃质齿冠和基腔部位,却具有更轻的δ 18O值,推测成岩作用对其改造程度却更显著[98]。因此,如何精确评估牙形刺氧同位素组成遭受成岩作用的改造程度,并优选氧同位素测试对象,有待进一步深入探讨。采用多种手段综合评估牙形刺不同部位的成岩作用改造程度,有助于有针对性地提取初始氧同位素信号,提升古温度重建可靠性。
-
牙形刺氧同位素古海水温度重建,必须考虑量化计算所带来的不确定性。磷酸盐氧同位素—温度经验公式的适用性、磷酸盐标样标定值的准确性、同时期海水氧同位素赋值的合理性等因素,均可能影响海水温度的定量重建结果,以及相关数据的对比分析及其地质意义解读。
目前,针对相同的牙形刺氧同位素值,基于不同的氧同位素—温度经验公式,重建的温度结果可呈现明显的偏差(表2)。事实上,相关氧同位素—温度经验公式各自基于不同种类的生物组织或器官(如藤壶、舌形贝和鲨鱼牙齿等)、不同的化学处理流程和磷酸盐标样而建立,它们是否、以及在多大程度上适用于已经绝灭的牙形动物,需要进一步探讨或优化。同时,由于不同生物生存的温度适宜区间有所不同,相关经验公式理论上也受限于一定的温度适用范围。目前,磷酸盐氧同位素—温度经验公式仍在不断优化改进[32-33,37,104],至今尚未达成共识。近年,Lécuyer et al.[37]通过将舌形贝和鲨鱼牙齿磷酸盐和同时期碳酸盐的氧同位素温度重建结果进行对比和校正,建立了新的磷酸盐氧同位素—温度经验公式(表2),得到广泛认可和应用[105-106]。此外,由于测试条件和流程差异,不同实验室对于国际磷灰石标样佛罗里达磷块岩氧同位素组成δ 18ONBS120c的测试结果存在可达0.9‰(VSMOW)的偏差[34-35,59]。这一定程度上也会影响相关测试结果,进而影响古海水温度量化计算结果及实验室间数据的对比讨论。对此,Pucéat et al.[33]将磷酸盐标样NBS120c的δ 18O测量值作为浮动变量纳入氧同位素—温度经验公式(表2),以弱化不同实验室测试偏差对温度重建结果的影响。
另一方面,海水δ 18Ow值是基于磷酸盐氧同位素—温度经验公式(表2)定量重建古海水温度的关键参数之一[15],但是其受控于多种影响因素而变化。目前普遍认为,由于冰川相对海水富集16O,全球海水平均δ 18Ow值主要受控于全球冰量体积的变化[2]:在冰期时偏高,间冰期时则偏低[15],呈现较好的冰期和间冰期旋回波动。同时,显生宙海水δ 18Ow值被认为受到海底热液活动的调控,长期稳定在0±1‰(VSMOW)[107-108],并呈现一定的长期演化[109]。Veizer和Prokoph et al.[4]基于对58 532个低镁方解石质生物壳体δ 18O值的统计分析认为,显生宙海水氧同位素组成以约+0.01‰/Ma(VSMOW)的速率不断变化。另一方面,局地海水δ 18Ow值如果受区域气候条件、水体循环特征等因素的影响,可以偏离全球海水平均值,呈现一定的空间变化[110]。例如,基于现代全球表层海水(水深<5 m)年平均氧同位素组成的观测结果表明,由于存在蒸发量/降水量比值差异,赤道地区海水δ 18Ow值相对极地地区偏重超过5‰,存在明显的纬度差异;同时,局限水体的δ 18Ow值相对开阔大洋可明显偏高[111-112]。因此,基于经验公式定量重建深时海水温度绝对值,其实是局部环境与全球气候背景的叠加信号[4,15],需综合考虑各种因素才能给予合理的地质解释(详见本文第5部分)。实践中,通常将δ 18Ow值简化为全球海水平均值并进行特定赋值,对于地球两极无冰时期(如早奥陶世)常假定为-1‰(VSMOW)[35],对于地球两极有冰时期(如二叠纪乌拉尔世)常假定为+1‰(VSMOW)[58]。
-
牙形刺氧同位素古温度计的应用虽然受限于多种因素的影响,但是对于重建深时海水温度仍具有不可替代的优势,可为我们理解古海洋、古气候、古地理和古生物演化提供宝贵信息,并已取得了大量研究成果。
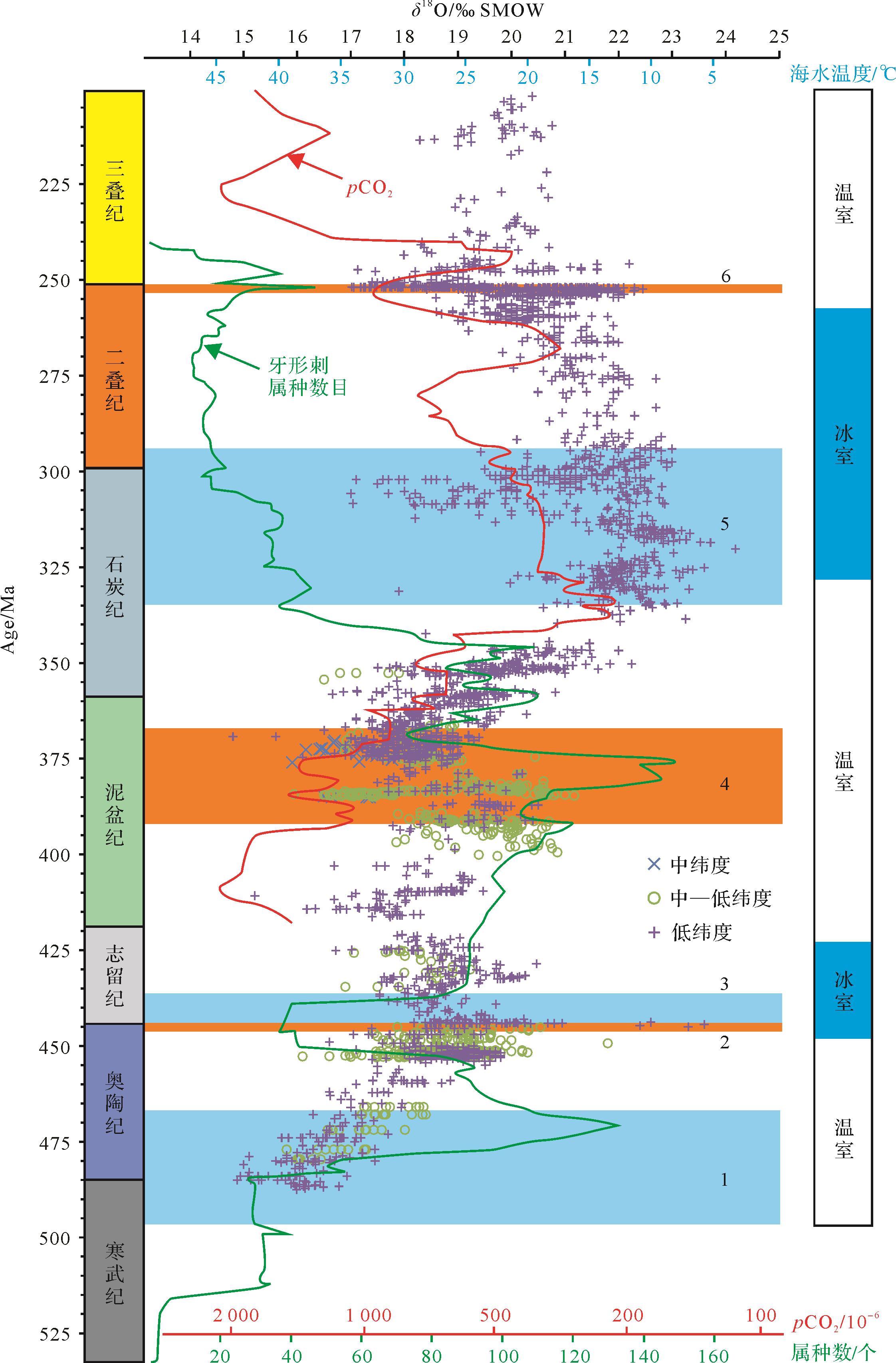
牙形刺氧同位素古温度重建,大致勾画出了晚寒武世至三叠纪海水温度演化曲线,重点揭示了关键地质历史时期海水温度特征,促进了对深时环境和生命演化的认识(图3)。例如,奥陶纪劳伦和冈瓦纳大陆牙形刺氧同位素证据显示:早奥陶世初期海水温度超过40 ℃,然后逐渐降低,至中奥陶世时与现今赤道海水的温度范围(28 ℃~32 ℃)相当,修正了奥陶纪总体处于超级温室状态的传统观点[35]。泥盆纪牙形刺氧同位素证据显示:中泥盆世温室气候传统观点难以成立,反之具有适宜的海水温度(23 ℃~25 ℃),并促进了生物礁的繁盛[60];晚泥盆世弗拉期—法门期(Frasnian-Famennian)转折时期全球气候变冷,低纬度海域降温5 ℃~7 ℃,生物面临严峻生存压力并走向了大绝灭[31,90]。晚石炭世北美中大陆地区牙形刺氧同位素研究揭示,同一准层序内牙形刺δ 18O差值可达1.7‰(VSMOW),可能受控于冰川成因的全球海平面变化,一定程度上可对比更新世冰期-间冰期情形[59]。早二叠世华南地区牙形刺氧同位素证据显示,晚古生代冰期的结束发生在空谷期,较传统观点(晚萨克马尔期)推迟了约10 Ma[58]。二叠纪—三叠纪转折时期华南和伊朗地区高分辨率牙形刺氧同位素证据显示,全球气候变暖滞后于该时期的生物大绝灭事件[40,115],更可能是早三叠世生物迟缓复苏的原因[24]。目前,牙形刺氧同位素古温度重建的相关研究主要集中于关键地质历史时期,而其他时期则相对薄弱甚至缺失,有待跟进;部分地质时期(如晚奥陶世和晚石炭世)牙形刺氧同位素所重建的古海水温度分歧较大,有时存在极值(如早奥陶世和早三叠世)(图3),其可靠性仍需进一步验证等。总体而言,牙形刺氧同位素古海水温度重建结果与前人基于其他指标重建的大气二氧化碳浓度曲线总体上具有相似的变化趋势[114](图3),进一步证实了该古温度计的有效性和合理性。

图 3 寒武纪至三叠纪牙形刺氧同位素及其定量重建的海水温度演化序列和环境—生物演化背景
Figure 3. The distribution of conodont oxygen isotopes and calculated paleotemperatures from the Cambrian to the Triassic and associated major environmental and biological events
另一方面,牙形刺氧同位素古温度重建,促进了对牙形动物生物习性的认识。例如,基于早奥陶世弗洛期劳伦大陆斜坡—盆地相牙形刺的氧同位素研究发现,不同属种牙形刺的δ 18O值存在1.6‰~1.8‰(VSMOW)的差异,被认为源于在海水温跃层背景下牙形动物的生物习性差异,并证明牙形动物以游泳分层而非底栖游移的方式生活[85]。基于泥盆纪弗拉期—法门期(Frasnian-Famennian)转折时期华南地区常见牙形动物的氧同位素研究发现,Polygnathus和Icriodus属牙形动物生活的水深范围较大,而Palmatolepis属牙形动物生活在表层水体中,是重建表层海水温度的优选对象[90]。基于石炭纪Donets盆地牙形刺氧同位素的研究发现,Gondolella属和Idioprioniodus属牙形刺的氧同位素值相对于同时期生活在表层水体的属种,在宾夕法尼亚亚纪早期较一致,在宾夕法尼亚亚纪晚期则偏高0.5‰~0.8‰(VSMOW),推测它们随着海侵过程中水体循环样式和初级生产力的变化而主动调整了生活水深[23]。基于二叠纪不同板块牙形刺的氧同位素研究发现,Streptognathodus属和Hindeodus属牙形刺主要生活于近表层海水环境,然而Jinogondolella属和Clarkina属牙形刺则可能在海水表层和深水环境中都能生存[58]。基于晚三叠世意大利Lagonegro盆地牙形刺的研究发现,Epigondolella triangularis种牙形刺氧同位素可合理对应同时期热带—亚热带海表温度范围,表明其属于海洋表层透光带内游泳生活方式[84]。因此,牙形刺氧同位素相关研究,通过重建特定属种牙形动物生活的海水温度,有效揭示了其生物习性与环境响应,具有重要应用价值。
此外,牙形刺氧同位素古温度重建,为古地理格局提供了关键约束。例如,基于奥陶纪阿根廷Precordillera地块高分辨率牙形刺氧同位素古温度重建结果,对比发现该地块与劳伦大陆所记录的古海水温度变化样式在早—中奥陶世基本一致,至晚奥陶世早期则存在约6 ℃的偏差,表明二者古地理格局发生了一定的演化[113]。通过三叠纪卡尼期-诺利期(Carnian-Norian)转折时期牙形刺氧同位素重建结果,对比发现Wrangellia地块和Pangea大陆该时期海水温度偏差约3 ℃~5 ℃,并结合古地磁数据可将前者古地理位置约束在约10°~15° N[116]。因此,牙形刺氧同位素相关研究,通过重建和对比不同地块的古海水温度特征及变化样式,可揭示它们之间的古地理演化关系,并有助于结合古地磁证据共同精准约束古地理位置。
-
牙形刺氧同位素古温度计由于具有定量重建古海水温度的优势,具有重要发展潜力;但是其地质意义的合理解读,受制于牙形动物自身生物习性、成岩作用改造程度、量化计算方式、古地理背景等因素,在应用时需加以关注和考量。例如,早三叠世同一层位不同属种牙形刺δ 18O值差异,响应了牙形动物属种差异[24];二叠纪—三叠纪转折时期扬子板块台地相至斜坡相不同剖面牙形刺δ 18O值差异,源于成岩作用不同程度的改造[40];石炭纪乌克兰半局限盆地相相对于中国华南地区斜坡相(纳庆剖面)牙形刺δ 18O值存在的显著差异(1‰~6‰),源于区域性季风环流的影响[23,117];二叠纪不同板块牙形刺δ 18O值差异,可能响应了各自蒸发量/降水量比值的不同[58]。
同时,牙形刺氧同位素古海水温度重建结果的对比与解析,需考虑古纬度、古地理和古海洋因素(图3)。例如,早—中奥陶世牙形刺氧同位素重建古温度显示,中—低纬度地区的海水温度低于低纬度地区[35,113,118],具有显著的温度—纬度分布特征。此外,Alberti et al.[119]在对中侏罗世腕足动物和双壳类壳体氧同位素研究的过程中,将海水δ 18Ow值分别进行-1‰(VSMOW)的传统赋值和参考温度—纬度梯度因素的改进赋值,发现基于后者所重建的古海水温度更加真实合理。另一方面,上升洋流发育的海域,如果深部不同温度和氧同位素特征的水体运移至陆架地区表层水体,可导致牙形刺氧同位素古温度重建结果具有明显的地域特色[120]。此外,局部水团的物理化学性质(如营养水平、浑浊度及氧气含量)也会影响牙形动物的生态分布[121],进而为牙形刺氧同位素古温度数据的地质意义解析带来一定的复杂性。
综上所述,基于有限研究区、特定属种牙形刺的氧同位素数据解读局部海水温度信号时需要十分谨慎。在确保牙形刺保存情况较好的前提下,不仅要考虑牙形动物自身生物习性对牙形刺氧同位素变化趋势的影响,也需要考虑研究区的古地理特征,包括局部气候条件(如盆地水体的局限性、蒸发量/降水量比值、季风气候等)、古纬度位置、洋流特征等因素对局部水体温度和氧同位素组成的影响,以正确对比和合理解读局部温度信号、及其与全球气候的关系。
-
牙形刺能较好的记录和保存原始海水氧同位素信号,同时,多年的机理探讨和分析测试方法开发使其氧同位素逐渐成为重建深时海水温度的关键温度计。目前,对牙形刺氧同位素古海水温度精准重建和精确解读,需综合考量生物习性、成岩作用、量化计算、古地理背景等多种因素的影响。该古温度计的广泛应用,极大地促进了我们对深时环境、海洋和生命演化的认识。未来,随着牙形刺氧同位素与海水温度关系的深入研究,分析测试方法的不断改进,其他古温度指标(如团簇同位素)的综合应用,以及对地质背景的全面理解,牙形刺氧同位素古温度计将在深时海水温度重建中发挥更大的作用。
Oxygen Isotope Palaeothermometry of Conodont Apatite: A review
-
摘要:
牙形刺是寒武系至三叠系海相地层里常见的磷酸盐质古生物化石,具有分布较广、易于保存等优势,其氧同位素可用于重建同时期海水温度,受到广泛关注和重视。从牙形刺基本特征、氧同位素古温度计原理、氧同位素分析测试方法等方面,对牙形刺氧同位素古温度计的基本特点和原理进行了回顾。在此基础上,进一步讨论了牙形动物生物习性、成岩作用改造、量化计算方式等多种因素对牙形刺氧同位素定量重建古海水温度的潜在影响。目前,牙形刺氧同位素古温度计在古海水温度重建、牙形动物生物习性研究、及古地理格局重建等方面开展了大量重要应用,相关研究成果极大促进了我们对深时环境和生物演化的认识。未来,深入研究牙形刺磷酸盐氧同位素与同时期海水的分馏机理、精细厘定牙形动物关键属种的生物习性、有效评估成岩作用对牙形刺氧同位素组成改造程度,并充分考量特定时期研究区古地理背景,将有助于牙形刺氧同位素古温度计在古海洋、古环境和古生物研究中发挥更大的作用。 Abstract:Conodonts are phosphatic fossils widely distributed in Cambrian to Triassic marine strata. Due to the advantage of well preservation, the oxygen isotope values of conodont apatite have been used to constrain coeval seawater temperature. In this context, we make a review of the development of oxygen isotope palaeothermometry, including the aspects of biological features, geochemical fundamentals, and oxygen isotope measuring methods of conodont apatite. We further summarize the three main factors that are critical for a quantitatively accurate reconstruction of the paleotemperature in deep time, including the life habit of conodont animals, diagenetic alteration of oxygen isotopes, and empirical formulas for paleotemperature value calculation based on phosphate oxygen isotopes. Presently, oxygen isotope palaeothermometry of conodont apatite has been widely and successfully applied in reconstructions of the paleo-oceanic temperature, living habit of conodont animals, and paleogeographic configurations, which have significantly enhanced our understanding of deep-time paleoenvironmental and life evolution. In the future, the development and application of this paleothermometer would be favored for more deep studies on the oxygen isotope fractionation mechanism between conodont and coeval seawater, the ecology of key conodont animal species, evaluation of diagenetic alteration, and full consideration of the paleogeographic background. Undoubtedly, oxygen isotope palaeothermometry of conodont apatite will play an indispensable role in paleo-oceanic, paleo-environmental, and paleontological studies. -
Key words:
- conodont /
- oxygen isotope /
- paleo-ocean /
- paleotemperature
-
图 1 牙形刺的形态和结构特征
(a,c)臀板刺属,(b,d)镰刺属,样品来自塔里木盆地柯坪地区奥陶系碳酸盐岩。图中A指示瓷白质齿冠,H指示玻璃质齿冠,B指示基腔
Figure 1. Morphological and structural features of conodonts
(a, c) Pygodus sp., (b, d) Drepanodus sp. conodonts, which were collected from Ordovician marine carbonates in the Kalpin area, Tarim Basin. A. albid crown; H. hyaline crown; B. basal body
图 2 基于TC⁃EA⁃IRMS和SIMS方法的牙形刺氧同位素分析测试流程示意图
Figure 2. Analytical workflow diagram of conodont oxygen isotopes through thermal conversion⁃elemental analyzer⁃isotope ratio mass spectrometry (TC⁃EA⁃IRMS) and secondary ion mass spectrometry (SIMS) methods
图 3 寒武纪至三叠纪牙形刺氧同位素及其定量重建的海水温度演化序列和环境—生物演化背景
牙形刺氧同位素数据整理自Huang et al.[90];Song et al.[105];Albanesi et al.[113]。CO2浓度重建结果和温/冰室气候划分据Foster et al.[114]修改。古温度的计算据Kolodny et al.[32]。假设温室气候下海水δ 18O平均值为-1‰(VSMOW),冰室气候时则为+1‰(VSMOW)。牙形刺属种数目变化曲线和重大生物演化事件据Fan et al.[49],其中1.奥陶纪生物大幅射;2.晚奥陶世生物大绝灭;3.早志留世生物辐射;4.中—晚泥盆世生物多样性骤减;5.晚石炭世—早二叠世生物辐射;6.二叠纪末期生物大绝灭。古纬度划分据Veizer et al.[4],分为中纬度(58°~35°)、中—低纬度(35°~26°)和低纬度(26°~0°)
Figure 3. The distribution of conodont oxygen isotopes and calculated paleotemperatures from the Cambrian to the Triassic and associated major environmental and biological events
Oxygen isotope data of conodont apatite compiled after Huang et al.[90]; Song et al.[105]; and Albanesi et al.[113]. CO2 concentration reconstruction and green/icehouse climate identification after Foster et al.[114]. Paleotemperature calculation is based on Kolodny et al.[32], assuming the seawater average δ 18O value of -1‰ (VSMOW) and +1‰ (VSMOW) for greenhouse and icehouse climates, respectively. Conodont species diversity curve and major bio⁃events are according to Fan et al.[49]: 1. Great Ordovician Biodiversification Event; 2. End⁃Ordovician mass extinction; 3. Early Silurian radiation; 4. Middle to Late Devonian diversity decline; 5. Late Carboniferous⁃Early Permian biodiversification; 6. End-Permian mass extinction. Paleolatitude is classified as middle latitude (58°-35°), mid⁃low latitude (35°-26°), and low latitude (26°-0°) according to Veizer et al.[4]
表 1 牙形刺的基本特征
Table 1.
The main characteristics of conodonts 成分 Ca5Na0.14(PO4)3.01(CO3)0.16F0.73(H2O)0.85 矿物 碳氟磷灰石 颜色 透明琥珀色、浅黄色或灰黑色等 形貌类型 单锥型、复合型和平台型 结构特征 基腔、齿冠(主齿) 寄主生物 底栖或分层游泳动物 时代分布 早寒武世—晚三叠世 生活环境 浅海陆架—深水盆地  下载: 导出CSV
下载: 导出CSV
表 2 常用的磷酸盐氧同位素—温度经验公式
注: 公式中T代表生物生存时环境水体的平均温度,单位为℃。δ 18Ophosphate代表生物矿化成因磷酸盐矿物的氧同位素组成。δ 18Ow代表海水的平均氧同位素组成,对于地球无极地冰川时通常假设为-1‰,有极地冰川时假设为+1‰[35,58]。δ 18ONBS120c代表国际磷灰石标样佛罗里达磷块岩的氧同位素组成,但是不同实验室报道的测量值有22.6‰[34]、22.4‰[59⁃60]、21.7‰[35]等,差值可达0.9‰。氧同位素数据均相对于VSMOW(维也纳标准平均大洋水)标准。Table 2.
Empirical formulas for paleotemperature reconstruction based on phosphate oxygen isotopes 温度计算经验公式 温度范围/℃ 研究对象 参考文献 T(℃)=111.4-4.3(δ 18Ophosphate-δ 18Ow) 3.5~27.3 藤壶和软体动物壳中的磷酸盐 Longinelli et al.[56]、Grossman[15] T(℃)=113.3-4.38(δ 18Ophosphate-δ 18Ow) 3.5~25 磷酸盐质鱼齿和鱼骨 Kolodny et al.[32]、Grossman[15] T(℃)=118.7-4.22[δ 18Ophosphate+(22.6-δ 18ONBS120c)-δ 18Ow] 3.5~28 磷酸盐质鱼齿 Pucéat et al.[33]、Grossman[15] T(℃)=117.4(±9.5)-4.50(±0.43)(δ 18Ophosphate-δ 18Ow) — 舌形贝和鲨鱼牙齿中的磷酸盐 Lécuyer et al.[37]
下载: 导出CSV
-
[1] Scotese C R, Song H J, Mills B J W, et al. Phanerozoic paleotemperatures: The earth’s changing climate during the last 540 million years[J]. Earth-Science Reviews, 2021, 215: 103503, doi: 10.1016/j.earscirev.2021.103503 . [2] Zachos J, Pagani M, Sloan L, et al. Trends, rhythms, and aberrations in global climate 65 Ma to present[J]. Science, 2001, 292(5517): 686-693. [3] Westerhold T, Marwan N, Drury A J, et al. An astronomically dated record of Earth’s climate and its predictability over the last 66 million years[J]. Science, 2020, 369(6509): 1383-1387. [4] Veizer J, Prokoph A. Temperatures and oxygen isotopic composition of Phanerozoic oceans[J]. Earth-Science Reviews, 2015, 146: 92-104. [5] Finnegan S, Bergmann K, Eiler J M, et al. The magnitude and duration of Late Ordovician–Early Silurian glaciation[J]. Science, 2011, 331(6019): 903-906. [6] Affek H P. Clumped isotope paleothermometry: Principles, applications, and challenges[J]. The Paleontological Society Papers, 2012, 18: 101-114. [7] Henkes G A, Passey B H, Grossman E L, et al. Temperature evolution and the oxygen isotope composition of Phanerozoic oceans from carbonate clumped isotope thermometry[J]. Earth and Planetary Science Letters, 2018, 490: 40-50. [8] Elderfield H, Ganssen G. Past temperature and δ 18O of surface ocean waters inferred from foraminiferal Mg/Ca ratios[J]. Nature, 2000, 405(6785): 442-445. [9] Jenkyns H C, Forster A, Schouten S, et al. High temperatures in the Late Cretaceous Arctic Ocean[J]. Nature, 2004, 432(7019): 888-892. [10] Mutterlose J, Malkoc M, Schouten S, et al. TEX86 and stable δ 18O paleothermometry of early Cretaceous sediments: Implications for belemnite ecology and paleotemperature proxy application[J]. Earth and Planetary Science Letters, 2010, 298(3/4): 286-298. [11] Prahl F G, Mix A C, Sparrow M A. Alkenone paleothermometry: Biological lessons from marine sediment records off western South America[J]. Geochimica et Cosmochimica Acta, 2006, 70(1): 101-117. [12] Łącka M, Cao M, Rosell-Melé A, et al. Postglacial paleoceanography of the western Barents Sea: Implications for alkenone-based sea surface temperatures and primary productivity[J]. Quaternary Science Reviews, 2019, 224: 105973, doi: 10.1016/j.quascirev.2019.105973 . [13] Veizer J, Ala D, Azmy K, et al. 87Sr/86Sr, δ 13C and δ 18O evolution of Phanerozoic seawater[J]. Chemical Geology, 1999, 161(1/2/3): 59-88. [14] Prokoph A, Shields G A, Veizer J. Compilation and time-series analysis of a marine carbonate δ 18O, δ 13C, 87Sr/86Sr and δ 34S database through Earth history[J]. Earth-Science Reviews, 2008, 87(3/4): 113-133. [15] Grossman E L. Oxygen isotope stratigraphy[M]//Gradstein F M, Ogg J G, Schmitz M D, et al. The geological time scale 2012. Amsterdam: Elsevier, 2012: 181-206. [16] Bennett C E, Williams M, Leng M J, et al. Oxygen isotope analysis of the eyes of pelagic trilobites: Testing the application of sea temperature proxies for the Ordovician[J]. Gondwana Research, 2018, 57: 157-169. [17] Blake R E, O’Neil J R, Garcia G A. Oxygen isotope systematics of biologically mediated reactions of phosphate: I. Microbial degradation of organophosphorus compounds[J]. Geochimica et Cosmochimica Acta, 1997, 61(20): 4411-4422. [18] Buggisch W, Joachimski M M, Sevastopulo G, et al. Mississippian δ 13Ccarb and conodont apatite δ 18O records: Their relation to the Late Palaeozoic Glaciation[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2008, 268(3/4): 273-292. [19] Rosenau N A, Herrmann A D, Leslie S A. Conodont apatite δ18O values from a platform margin setting, Oklahoma, USA: Implications for initiation of Late Ordovician icehouse conditions[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2012, 315-316: 172-180. [20] Chen B, Joachimski M M, Sun Y D, et al. Carbon and conodont apatite oxygen isotope records of Guadalupian–Lopingian boundary sections: Climatic or sea-level signal?[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2011, 311(3/4): 145-153. [21] Trotter J A, Williams I S, Nicora A, et al. Long-term cycles of Triassic climate change: A new δ 18O record from conodont apatite[J]. Earth and Planetary Science Letters, 2015, 415: 165-174. [22] Bergmann K D, Finnegan S, Creel R, et al. A paired apatite and calcite clumped isotope thermometry approach to estimating Cambro-Ordovician seawater temperatures and isotopic composition[J]. Geochimica et Cosmochimica Acta, 2018, 224: 18-41. [23] Montañez I P, Osleger D J, Chen J T, et al. Carboniferous climate teleconnections archived in coupled bioapatite δ 18OPO4 and 87Sr/86Sr records from the epicontinental Donets Basin, Ukraine[J]. Earth and Planetary Science Letters, 2018, 492: 89-101. [24] Sun Y D, Joachimski M M, Wignall P B, et al. Lethally hot temperatures during the Early Triassic greenhouse[J]. Science, 2012, 338(6105): 366-370. [25] Wang W Q, Garbelli C, Zhang F F, et al. A high-resolution Middle to Late Permian paleotemperature curve reconstructed using oxygen isotopes of well-preserved brachiopod shells[J]. Earth and Planetary Science Letters, 2020, 540: 116245, doi: 10.1016/j.epsl.2020.116245 . [26] Nicoll R S, Metcalfe I, Wang C Y. New species of the conodont Genus Hindeodus and the conodont biostratigraphy of the Permian-Triassic boundary interval[J]. Journal of Asian Earth Sciences, 2002, 20(6): 609-631. [27] 王志浩,祁玉平. 我国北方石炭—二叠系牙形刺序列再认识[J]. 微体古生物学报,2003,20(3):225-243. Wang Zhihao, Qi Yuping. Review of Carboniferous-Permian conodont biostratigraphy in North China[J]. Acta Micropalaeontologica Sinica, 2003, 20(3): 225-243. [28] Kaufmann B. Calibrating the Devonian Time Scale: A synthesis of U-Pb ID-TIMS ages and conodont stratigraphy[J]. Earth-Science Reviews, 2006, 76(3/4): 175-190. [29] Bergström S M, Ferretti A. Conodonts in Ordovician biostratigraphy[J]. Lethaia, 2017, 50(3): 424-439. [30] Wenzel B, Lécuyer C, Joachimski M M. Comparing oxygen isotope records of Silurian calcite and phosphate: δ18O compositions of brachiopods and conodonts[J]. Geochimica et Cosmochimica Acta, 2000, 64(11): 1859-1872. [31] Joachimski M M, Buggisch W. Conodont apatite δ18O signatures indicate climatic cooling as a trigger of the Late Devonian mass extinction[J]. Geology, 2002, 30(8): 711-714. [32] Kolodny Y, Luz B, Navon O. Oxygen isotope variations in phosphate of biogenic apatites, I. Fish bone apatite: Rechecking the rules of the game[J]. Earth and Planetary Science Letters, 1983, 64(3): 398-404. [33] Pucéat E, Joachimski M M, Bouilloux A, et al. Revised phosphate-water fractionation equation reassessing paleotemperatures derived from biogenic apatite[J]. Earth and Planetary Science Letters, 2010, 298(1/2): 135-142. [34] Vennemann T W, Fricke H C, Blake R E, et al. Oxygen isotope analysis of phosphates: A comparison of techniques for analysis of Ag3PO4 [J]. Chemical Geology, 2002, 185(3/4): 321-336. [35] Trotter J A, Williams I S, Barnes C R, et al. Did cooling oceans trigger Ordovician biodiversification? Evidence from conodont thermometry[J]. Science, 2008, 321(5888): 550-554. [36] Wheeley J R, Smith M P, Boomer I. Oxygen isotope variability in conodonts: Implications for reconstructing Palaeozoic palaeoclimates and palaeoceanography[J]. Journal of the Geological Society, 2012, 169(3): 239-250. [37] Lécuyer C, Amiot R, Touzeau A, et al. Calibration of the phosphate δ 18O thermometer with carbonate-water oxygen isotope fractionation equations[J]. Chemical Geology, 2013, 347: 217-226. [38] Trotter J A, Eggins S M. Chemical systematics of conodont apatite determined by laser ablation ICPMS[J]. Chemical Geology, 2006, 233(3/4): 196-216. [39] Trotter J A, Gerald J D F, Kokkonen H, et al. New insights into the ultrastructure, permeability, and integrity of conodont apatite determined by transmission electron microscopy[J]. Lethaia, 2007, 40(2): 97-110. [40] Chen J, Shen S Z, Li X H, et al. High-resolution SIMS oxygen isotope analysis on conodont apatite from South China and implications for the end-Permian mass extinction[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2016, 448: 26-38. [41] Pander C H. Monographie der fossilen fische des silurischen systems der Russisch-Baltischen gouvernements[M]. St. Petersburg: Buchdruckerei Kaiserlichen Akademie des Wissenschaften, 1856: 91. [42] Pietzner H, Vahl J, Werner H, et al. Zur chemischen zusammensetzung und mikromorphologie der conodonten[J]. Palaeontographica Abteilung A, 1968, 128(4/5/6): 115-152. [43] Wright J. Conodont apatite: Structure and geochemistry[J]. Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends, 1990, 1: 445-459. [44] Donoghue P C, Forey P L, Aldridge R J. Conodont affinity and chordate phylogeny[J]. Biological Reviews, 2000, 75(2): 191-251. [45] Sweet W C, Donoghue P C J. Conodonts: Past, present, future[J]. Journal of Paleontology, 2001, 75(6): 1174-1184. [46] Blieck A, Turner S, Burrow C J, et al. Fossils, histology, and phylogeny: Why conodonts are not vertebrates[J]. Episodes, 2010, 33(4): 234-241. [47] Murdock D J E, Dong X P, Repetski J E, et al. The origin of conodonts and of vertebrate mineralized skeletons[J]. Nature, 2013, 502(7472): 546-549. [48] Balter V, Martin J E, Tacail T, et al. Calcium stable isotopes place Devonian conodonts as first level consumers[J]. Geochemical Perspectives Letters, 2019, 10: 36-39. [49] Fan J X, Shen S Z, Erwin D H, et al. A high-resolution summary of Cambrian to Early Triassic marine invertebrate biodiversity[J]. Science, 2020, 367(6475): 272-277. [50] Sharp Z D, Atudorei V, Furrer H. The effect of diagenesis on oxygen isotope ratios of biogenic phosphates[J]. American Journal of Science, 2000, 300(3): 222-237. [51] Lecuyer C, Grandjean P, Sheppard S M F. Oxygen isotope exchange between dissolved phosphate and water at temperatures ≤135°C: Inorganic versus biological fractionations[J]. Geochimica et Cosmochimica Acta, 1999, 63(6): 855-862. [52] LeGeros R Z. Apatites in biological systems[J]. Progress in Crystal Growth and Characterization, 1981, 4(1/2): 1-45. [53] Hughes J M, Rakovan J. The crystal structure of apatite, Ca5(PO4)3(F, OH, Cl)[J]. Reviews in Mineralogy and Geochemistry, 2002, 48(1): 1-12. [54] Kemp A. Amino acid residues in conodont elements[J]. Journal of Paleontology, 2002, 76(3): 518-528. [55] Žigaitė Ž, Qvarnström M, Bancroft A, et al. Trace and rare earth element compositions of Silurian conodonts from the Vesiku Bone Bed: Histological and palaeoenvironmental implications[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 549: 109449, doi: 10.1016/j.palaeo.2019.109449 . [56] Longinelli A, Nuti S. Revised phosphate-water isotopic temperature scale[J]. Earth and Planetary Science Letters, 1973, 19(3): 373-376. [57] Urey H C, Lowenstam H A, Epstein S, et al. Measurement of paleotemperatures and temperatures of the Upper Cretaceous of England, Denmark, and the southeastern United States[J]. Geological Society of America Bulletin, 1951, 62(4): 399-416. [58] Chen B, Joachimski M M, Shen S Z, et al. Permian ice volume and palaeoclimate history: Oxygen isotope proxies revisited[J]. Gondwana Research, 2013, 24(1): 77-89. [59] Joachimski M M, von Bitter P H, Buggisch W. Constraints on Pennsylvanian glacioeustatic sea-level changes using oxygen isotopes of conodont apatite[J]. Geology, 2006, 34(4): 277-280. [60] Joachimski M M, Breisig S, Buggisch W, et al. Devonian climate and reef evolution: Insights from oxygen isotopes in apatite[J]. Earth and Planetary Science Letters, 2009, 284(3/4): 599-609. [61] Tudge A P. A method of analysis of oxygen isotopes in orthophosphate: Its use in the measurement of paleotemperatures[J]. Geochimica et Cosmochimica Acta, 1960, 18(1/2): 81-93. [62] Crowson R A, Showers W J, Wright E K, et al. Preparation of phosphate samples for oxygen isotope analysis[J]. Analytical Chemistry, 1991, 63(20): 2397-2400. [63] Stuart-Williams H L Q, Schwarcz H P. Oxygen isotopic analysis of silver orthophosphate using a reaction with bromine[J]. Geochimica et Cosmochimica Acta, 1995, 59(18): 3837-3841. [64] Holmden C, Papanastassiou D A, Wasserburg G J. Negative thermal ion mass spectrometry of oxygen in phosphates[J]. Geochimica et Cosmochimica Acta, 1997, 61(11): 2253-2263. [65] O’Neil J R, Roe L J, Reinhard E, et al. A rapid and precise method of oxygen isotope analysis of biogenic phosphate[J]. Israel Journal of Earth Sciences, 1994, 43(3): 203-212. [66] Laporte D F, Holmden C, Patterson W P, et al. Oxygen isotope analysis of phosphate: Improved precision using TC/EA CF‐IRMS[J]. Journal of Mass Spectrometry, 2009, 44(6): 879-890. [67] Grimes V, Pellegrini M. A comparison of pretreatment methods for the analysis of phosphate oxygen isotope ratios in bioapatite[J]. Rapid Communications in Mass Spectrometry, 2013, 27(3): 375-390. [68] Griffin J M, Montañez I P, Matthews J A, et al. A refined protocol for δ 18OPO 4 analysis of conodont bioapatite[J]. Chemical Geology, 2015, 417: 11-20. [69] Cid-Andres A P. A review on useful concepts for stable isotope of oxygen in phosphate (δ 18Op) extraction, purification and analysis of freshwater samples and other potential phosphate sources[J]. Microchemical Journal, 2015, 123: 105-110. [70] 杜勇,朱园园,宋虎跃,等. 微量磷灰石中磷酸根氧同位素分析方法[J]. 地球科学,2019,44(2):456-462. Du Yong, Zhu Yuanyuan, Song Huyue, et al. Analytical method for δ 18O of phosphate in trace apatite[J]. Earth Science, 2019, 44(2): 456-462. [71] Joachimski M M, Lai X L, Shen S Z, et al. Climate warming in the latest Permian and the Permian-Triassic mass extinction[J]. Geology, 2012, 40(3): 195-198. [72] Schobben M, Joachimski M M, Korn D, et al. Palaeotethys seawater temperature rise and an intensified hydrological cycle following the end-Permian mass extinction[J]. Gondwana Research, 2014, 26(2): 675-683. [73] Trotter J A, Williams I S, Barnes C R, et al. New conodont δ 18O records of Silurian climate change: Implications for environmental and biological events[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2016, 443: 34-48. [74] Stephan E. Oxygen isotope analysis of animal bone phosphate: Method refinement, influence of consolidants, and reconstruction of palaeotemperatures for Holocene sites[J]. Journal of Archaeological Science, 2000, 27(6): 523-535. [75] Pederzani S, Snoeck C, Wacker U, et al. Anion exchange resin and slow precipitation preclude the need for pretreatments in silver phosphate preparation for oxygen isotope analysis of bioapatites[J]. Chemical Geology, 2020, 534: 119455, doi: 10.1016/j.chemgeo.2019.119455 . [76] 高建飞,丁悌平. 激光熔蚀微量氧同位素分析方法及其地质应用[J]. 地质论评,2008,54(1):139-144. Gao Jianfei, Ding Tiping. Laser microprobe oxygen isotope analysis method and geology applications[J]. Geological Review, 2008, 54(1): 139-144. [77] 王润,陈剑波,赵来时,等. 二次离子质谱微区原位牙形石氧同位素分析及其在古海表水温记录中的应用[J]. 世界地质,2013,32(4):652-658. Wang Run, Chen Jianbo, Zhao Laishi, et al. In situ oxygen isotope analysis of conodonts by SIMS and its implication for paleo-sea surface temperature[J]. Global Geology, 2013, 32(4): 652-658. [78] Aubert M, Williams I S, Boljkovac K, et al. In situ oxygen isotope micro-analysis of faunal material and human teeth using a SHRIMP II: A new tool for palaeo-ecology and archaeology[J]. Journal of Archaeological Science, 2012, 39(10): 3184-3194. [79] 周丽芹, Williams I S,刘建辉,等. 牙形石SHRIMP微区原位氧同位素分析方法[J]. 地质学报,2012,86(4):611-618. Zhou Liqin, Williams I S, Liu Jianhui, et al. Methodology of SHRIMP in-situ o isotopes analysis on conodont[J]. Acta Geologica Sinica, 2012, 86(4): 611-618. [80] Li X H, Li W X, Wang X C, et al. Role of mantle-derived magma in genesis of early Yanshanian granites in the Nanling Range, South China: In situ zircon Hf-O isotopic constraints[J]. Science in China Series D: Earth Sciences, 2009, 52(9): 1262-1278. [81] Li X H, Long W G, Li Q L, et al. Penglai zircon megacrysts: A potential new working reference material for microbeam determination of Hf–O isotopes and U–Pb age[J]. Geostandards and Geoanalytical Research, 2010, 34(2): 117-134. [82] Rigo M, Trotter J A, Preto N, et al. Oxygen isotopic evidence for Late Triassic monsoonal upwelling in the northwestern Tethys[J]. Geology, 2012, 40(6): 515-518. [83] Sun Y D, Wiedenbeck M, Joachimski M M, et al. Chemical and oxygen isotope composition of gem-quality apatites: Implications for oxygen isotope reference materials for secondary ion mass spectrometry (SIMS)[J]. Chemical Geology, 2016, 440: 164-178. [84] Rigo M, Joachimski M M. Palaeoecology of Late Triassic conodonts: Constraints from oxygen isotopes in biogenic apatite[J]. Acta Palaeontologica Polonica, 2010, 55(3): 471-478. [85] Wheeley J R, Jardine P E, Raine R J, et al. Paleoecologic and paleoceanographic interpretation of δ18O variability in Lower Ordovician conodont species[J]. Geology, 2018, 46(5): 467-470. [86] Clark D L. Conodont biofacies and provincialism[M]. Geological Society of America Special Paper 196, 1984: 1-340. [87] Rasmussen J A, Stouge S. Baltoscandian conodont biofacies fluctuations and their link to Middle Ordovician (Darriwilian) global cooling[J]. Palaeontology, 2018, 61(3): 391-416. [88] Barnes C R, Fahreus L E. Provinces, communities, and the proposed nektobenthic habit of Ordovician conodontophorids[J]. Lethaia, 1975, 8(2): 133-149. [89] Seddon G, Sweet W C. An ecologic model for conodonts[J]. Journal of Paleontology, 1971, 45(5): 869-880. [90] Huang C, Joachimski M M, Gong Y M. Did climate changes trigger the Late Devonian Kellwasser Crisis? Evidence from a high-resolution conodont δ 18OPO4 record from South China[J]. Earth and Planetary Science Letters, 2018, 495: 174-184. [91] Sweet W C. The conodonta: Morphology, taxonomy, paleoecology, and evolutionary history of a long-extinct animal phylum[M]. Oxford: Clarendon Press, 1988. [92] Buggisch W, Joachimski M M, Lehnert O, et al. Did intense volcanism trigger the first Late Ordovician icehouse?[J]. Geology, 2010, 38(4): 327-330. [93] Quinton P C, Macleod K G. Oxygen isotopes from conodont apatite of the midcontinent, US: Implications for Late Ordovician climate evolution[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 404: 57-66. [94] MacLeod K G. Conodonts and the paleoclimatological and paleoecological applications of phosphate δ 18O measurements[J]. The Paleontological Society Papers, 2012, 18: 69-84. [95] Barham M, Joachimski M M, Murray J, et al. Diagenetic alteration of the structure and δ 18O signature of Palaeozoic fish and conodont apatite: Potential use for corrected isotope signatures in palaeoenvironmental interpretation[J]. Chemical Geology, 2012, 298-299: 11-19. [96] Epstein A G, Epstein J B, Harris L D. Conodont color alteration: An index to organic metamorphism[R]. Washington: United States Government Printing Office, 1977: 1-27. [97] Rejebian V A, Harris A G, Huebner J S. Conodont color and textural alteration: An index to regional metamorphism, contact metamorphism, and hydrothermal alteration[J]. Geological Society of America Bulletin, 1987, 99(4): 471-479. [98] Zhang L, Cao L, Zhao L S, et al. Raman spectral, elemental, crystallinity, and oxygen-isotope variations in conodont apatite during diagenesis[J]. Geochimica et Cosmochimica Acta, 2017, 210: 184-207. [99] Shemesh A. Crystallinity and diagenesis of sedimentary apatites[J]. Geochimica et Cosmochimica Acta, 1990, 54(9): 2433-2438. [100] Pucéat E, Reynard B, Lécuyer C. Can crystallinity be used to determine the degree of chemical alteration of biogenic apatites?[J]. Chemical Geology, 2004, 205(1/2): 83-97. [101] Habermann D, Götte T, Meijer J, et al. High resolution rare-earth elements analyses of natural apatite and its application in geo-sciences: Combined micro-PIXE, quantitative CL spectroscopy and electron spin resonance analyses[J]. Nuclear Instruments and Methods in Physics Research Section B: Beam Interactions with Materials and Atoms, 2000, 161-163: 846-851. [102] Golding M L, McMillan R. The impacts of diagenesis on the geochemical characteristics and Color Alteration Index of conodonts[J]. Palaeobiodiversity and Palaeoenvironments, 2020, doi: 10.1007/s12549-020-00447-y . [103] McMillan R, Golding M. Thermal maturity of carbonaceous material in conodonts and the Color Alteration Index: Independently identifying maximum temperature with Raman spectroscopy[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2019, 534: 109290, doi: 10.1016/j.palaeo.2019.109290 . [104] Chang S J, Blake R E. Precise calibration of equilibrium oxygen isotope fractionations between dissolved phosphate and water from 3 to 37 ℃[J]. Geochimica et Cosmochimica Acta, 2015, 150: 314-329. [105] Song H J, Wignall P B, Song H Y, et al. Seawater temperature and dissolved oxygen over the past 500 million years[J]. Journal of Earth Science, 2019, 30(2): 236-243. [106] Goudemand N, Romano C, Leu M, et al. Dynamic interplay between climate and marine biodiversity upheavals during the early Triassic Smithian-Spathian biotic crisis[J]. Earth-Science Reviews, 2019, 195: 169-178. [107] Ryb U, Eiler J M. Oxygen isotope composition of the Phanerozoic ocean and a possible solution to the dolomite problem[J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(26): 6602-6607. [108] Jaffrés J B D, Shields G A, Wallmann K. The oxygen isotope evolution of seawater: A critical review of a long-standing controversy and an improved geological water cycle model for the past 3.4 billion years[J]. Earth-Science Reviews, 2007, 83(1/2): 83-122. [109] Johnson B W, Wing B A. Limited Archaean continental emergence reflected in an early Archaean 18O-enriched ocean[J]. Nature Geoscience, 2020, 13(3): 243-248. [110] Macarewich S I, Poulsen C J, Montañez I P. Simulation of oxygen isotopes and circulation in a Late Carboniferous epicontinental sea with implications for proxy records[J]. Earth and Planetary Science Letters, 2021, 559: 116770, doi: 10.1016/j.epsl.2021.116770 . [111] LeGrande A N, Schmidt G A. Global gridded data set of the oxygen isotopic composition in seawater[J]. Geophysical Research Letters, 2006, 33(12): L12604, doi: 10.1029/2006GL026011 . [112] Rohling E J. Oxygen isotope composition of seawater[M]//Elias S A. The encyclopedia of Quaternary science. Amsterdam: Elsevier, 2013, 2: 915-922. [113] Albanesi G L, Barnes C R, Trotter J A, et al. Comparative Lower-Middle Ordovician conodont oxygen isotope palaeothermometry of the Argentine Precordillera and Laurentian margins[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 549: 109115, doi: 10.1016/j.palaeo.2019.03.016 . [114] Foster G L, Royer D L, Lunt D J. Future climate forcing potentially without precedent in the last 420 million years[J]. Nature Communications, 2017, 8: 14845. [115] Chen J, Shen S Z, Zhang Y C, et al. Abrupt warming in the latest Permian detected using high-resolution in situ oxygen isotopes of conodont apatite from Abadeh, central Iran[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 560: 109973, doi: 10.1016/j.palaeo.2020.109973 . [116] Sun Y D, Orchard M J, Kocsis Á T, et al. Carnian–Norian (Late Triassic) climate change: Evidence from conodont oxygen isotope thermometry with implications for reef development and Wrangellian tectonics[J]. Earth and Planetary Science Letters, 2020, 534: 116082, doi: 10.1016/j.epsl.2020.116082 . [117] Chen B, Joachimski M M, Wang X D, et al. Ice volume and paleoclimate history of the Late Paleozoic Ice Age from conodont apatite oxygen isotopes from Naqing (Guizhou, China)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2016, 448: 151-161. [118] Jin J S, Zhan R B, Wu R C. Equatorial cold-water tongue in the Late Ordovician[J]. Geology, 2018, 46(9): 759-762. [119] Alberti M, Leshno Y, Fürsich F T, et al. Stress in the tropics? Impact of a latitudinal seawater δ18O gradient on Middle Jurassic temperature reconstructions at low latitudes[J]. Geology, 2020, 48(12): 1210-1215. [120] Zhu J, Poulsen C J, Otto-Bliesner B L, et al. Simulation of early Eocene water isotopes using an Earth system model and its implication for past climate reconstruction[J]. Earth and Planetary Science Letters, 2020, 537: 116164, doi: 10.1016/j.epsl.2020.116164 . [121] Herrmann A D, Barrick J E, Algeo T J. The relationship of conodont biofacies to spatially variable water mass properties in the Late Pennsylvanian Midcontinent Sea[J]. Paleoceanography, 2015, 30(3): 269-283. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 486
- HTML全文浏览量: 146
- PDF下载量: 165
- 被引次数: 0

