

 下载:
下载:






-
生物扰动指生物在生命活动过程中对生存环境周围的沉积物颗粒所进行的搅动、混合和破坏,所形成的各类生物成因沉积构造[1],它是一种典型的小规模但潜在的重要地质过程,能改造或改变岩石的性质[2⁃3],比如岩石的孔隙度、渗透率和连通性[4]。这一过程会对原生沉积岩性造成破坏和蚀变,也是生物调节沉积物物理和化学特征的结果[5⁃6]。生物扰动可增强或减弱油气水储层质量及其流动特性[7]。目前,生物扰动对储集层物性存在不同程度的影响[8],一些学者研究发现生物扰动可以使砂泥混合物中的泥质含量降低,使其储集性能变好,从而提高油气的可采率[9];而另外一部分学者发现,生物扰动初始提高了储集层的孔隙度和渗透率,但在后期成岩过程中由于成岩流体的充注,产生多期次的胶结物,使得储集层的岩石物性又变差[9],并增加了储层的非均质性或沉积物的均质化[10],从而导致油气在运移过程中发生淤塞或沉淀。
目前,生物扰动储集层研究主要集中在储集空间和孔隙结构特征、储集层内流体的传输特征、影响生物扰动储集层渗透性的主控因素和生物扰动储集层类型划分及其属性特征等方面。在生物扰动储集层储集空间和孔隙结构研究方面,众多研究学者采用了多种不同的方法表征岩石孔隙结构,比如铸体薄片、扫描电镜(SEM)、场发射扫描电镜(FESEM)、聚焦离子束扫描电镜(FIB-SEM)、核磁共振(NMR)、压汞法(MIP)、低压气体吸附法(N2、CO2)、场发射扫描电子显微镜(FESEM)和能量色散X射线光谱(EDS)[11]等,这些方法可直接定性或定量分析岩石孔隙结构特征,如孔隙半径、孔隙几何形状、孔隙类型等,或者通过计算参数来定量描述岩石孔隙结构特征,如孔隙半径、孔径分布、孔喉大小、孔体积、比表面积、分形维数等[12]。但这些方法通常会对样品有损伤,或者由于各种人为因素和仪器的原因造成的误差可能比较大,或者无法获得非连通孔隙的参数[13⁃15]。近年来,微纳米CT(micro-CT、nano-CT)等新型无损检测技术被用来三维可视化表征生物扰动储集空间的内部结构[16⁃20],提高了相关资源量评估的准确性和推动制定出更合理的开发方案[21]。然而,上述相关研究以野外露头和岩心详细观测为基础,难免受到野外露头分布或井下取心数量的限制;因此,利用高精度电成像测井资料进行生物扰动储集层研究将可能成为最有潜力的手段之一[22]。
在综合分析前人关于生物扰动储层相关研究的基础上,分别从生物扰动碎屑岩和碳酸盐岩储层的储集空间类型、孔隙结构、储层介质类型、流体传输特征、储层类型及其属性特征等方面综述生物扰动储集层特征,这对了解这类非常规油气水储集层的研究进展,指导这类油气藏的远景勘探、储量计算以及选择合理的开发方案具有重要的借鉴意义[23]。
-
生物扰动已被证明可以改变沉积物的结构和结构属性[24⁃27]。在生物扰动碎屑岩储集层中,典型的储集空间主要为原生粒间孔、次生溶蚀孔和生物铸模孔等。例如Tonkin et al.[28]在对Jeanne d’arc盆地白垩系Ben Nevis组研究发现,富泥的细粒砂岩岩石矿物组成以石英为主,含少量的长石、方解石、岩屑、黏土矿物和有机碎屑,发育良好的原生粒间孔隙,这些原生粒间孔隙存于分选中等—良好、次角状—次圆状松散堆积的颗粒之间(图1,2),孔隙度为4.9%~27.2%。Ophiomorpha生物潜穴是整口井最显著的遗迹化石,Ophiomorpha生物潜穴降低了原始地层的孔隙度和渗透率,然而砂质充填的Thalassinoides生物潜穴被认为提高了岩石物性[28]。在其他生物扰动碎屑岩储层研究实例中发现,早期的原生孔隙因为经历了广泛的成岩溶蚀后再被方解石胶结[29⁃30],差异性充填方解石胶结物显著降低了储层的孔隙度和渗透率[31]。但若在随后的成岩过程中这些方解石胶结物又被溶解,则又可以使得这些粒间孔隙得以保留[28]。此外,生物扰动还可能通过分离物理分选的颗粒来增加各向同性,或者通过分选颗粒和创建潜穴系统来减少各向同性[28]。


在生物扰动碳酸盐岩储集层中,典型的储集空间为原生粒间孔、晶间孔和溶蚀孔等。如柴达木盆地南缘石炭系碳酸盐岩中的Chondrites潜穴多被方解石充填,而方解石质的潜穴充填物后期又多被溶蚀,形成了有效的储集空间[32]。生物潜穴充填物的孔隙类型还有白云石晶间孔、晶间次生溶孔和晶内次生溶孔[33⁃34]。白云岩填充的潜穴中存在硫酸盐还原细菌、还原条件和海洋有机物[35⁃36]。溶蚀作用是生物扰动储集层形成有效储集空间的重要作用[37⁃44],且同生期—早成岩期溶蚀作用是储集空间发育的关键因素;白云化作用也对储集性能具有积极意义[45⁃47]。众多研究表明储集空间差异化的保存,取决于埋藏胶结强度、白云化程度与烃类充注程度[48⁃49]。生物通常通过诸如沉积物压实、沉积物分选和向沉积物中添加胞外多糖(即EPS或黏液)等活动来改变基质的物理和化学组成[50]。
在不同的地质时期,伴随着生物演化和沉积环境变化的影响生物扰动构造也发生变化,总体形态上呈现由简到繁,分布范围由小到大。在寒武纪生命大爆发时期,开始出现具有垂向分量的生物潜穴;到奥陶纪生物大辐射时期,海相遗迹化石的分布得到扩展,逐渐从滨、浅海扩展至半深海和深海;而中生代海洋革命时期,海相和陆相遗迹化石同时增多,形态类型多样,生物对生态空间的利用也增强[51]。这些遗迹化石或生物扰动构造的变化又会改造原始地层的储集空间,进而在不同程度上影响地层的储集能力。
-
前人众多研究表明生物扰动可以影响储层的渗透率和孔隙度[10,52⁃53]。根据生物扰动储层的物性特征和流体传输特征。Pemberton et al.[54]和Gingras et al.[55⁃56]将生物扰动储层划分两种类型:双孔隙度生物成因储层介质和双渗透率生物成因储层介质。双孔隙度生物成因储层介质是指潜穴渗透率与基质渗透率之间的差异小于两个数量级的层段[55⁃56],生物扰动可以施加较小对比的渗透场。双渗透率生物成因储层介质是指基质渗透率与地层渗透率相差3个数量级以上的层段[55],生物扰动可以形成显著的、高度对比的渗透场。两种生物成因储层介质都影响含流体岩石的储层质量[56⁃57],它们可以存在于同一生物扰动层内,也可以发育在不同的区域[35]。
在双孔隙度生物成因储层介质中,岩石体积的大部分用于导流[56]。尽管通过高渗透率区域的流体通量较大,但生物潜穴与基质之间的流动相互作用非常广泛(图3)。这对储层行为有三点重要影响:(1)更大的孔隙体积有助于产液或产气;(2)由于毛细管效应,在多相流动中,不同流体相可能发生分离(油可能被分离在储层的细粒部分);(3)平流作用大于扩散作用(资源在地层中的流动响应于水力梯度)。Knaust et al.[58]对Gullfaks油田马斯特里赫特的分析表明,孔隙度在基质中提供了一个连通域,形成了开放的生物潜穴和裂缝,从而提高了油气产量[58]。一般来说,在双孔隙度生物成因储层介质中输送的流体不可能均匀运动,因此生物潜穴直径和弯曲度就显得很重要[59]。流体优先沿着高渗透率地层流动,其中流动路径根据生物潜穴形态可以变得弯曲[56]。

双渗透率生物成因储层介质指的是基质和裂缝相互连通的储层,因此两者都对流体流动有重要贡献[60]。在双渗透率生物成因储层介质中(图4),基质相对于生物潜穴具有较差的流体流动特性(尽管生物潜穴可能提供良好的气藏)[56]。双渗透率生物成因储层介质对储层和含水层的影响包括:(1)只有岩石的高渗透部分有助于流动;(2)基质主要通过扩散与流动管道相互作用;(3)高度对比的渗透场可能导致油藏“水窜”或降低二次采油技术(如水驱)的有效性[56,61]。另一方面,双渗透率生物成因储层介质的存在可能不影响产气。这是因为气体比液体更容易通过弯曲的网络流动,并且气体可以储存在基质中并通过渗透条纹输送。双渗透率生物成因储层介质是生物潜穴通过粗颗粒被动充填、粗粒物质主动回填或碳酸盐地层中与潜穴有关的成岩作用形成[56]。

关于生物扰动储层中流体的传输特征研究,目前主要有物理(实验)模拟和数值(实验)模拟两种研究思路。物理实验模拟的优点是岩样直接源于真实地层,给人感官上更可信;它的缺点是花费成本比较高,同时实验过程中,岩心分析技术会受到一定的限制(主要是因为暴露表面狭窄),并且涉及岩心时技术性质的表征比较复杂[62],由于各种人为因素和仪器的原因造成的误差可能比较大。而数值实验模拟的优点是成本相对低廉,人为和仪器原因造成的误差小;缺点是由于数据不是直接来源于真实地层,给人的感官没那么可靠[18]。
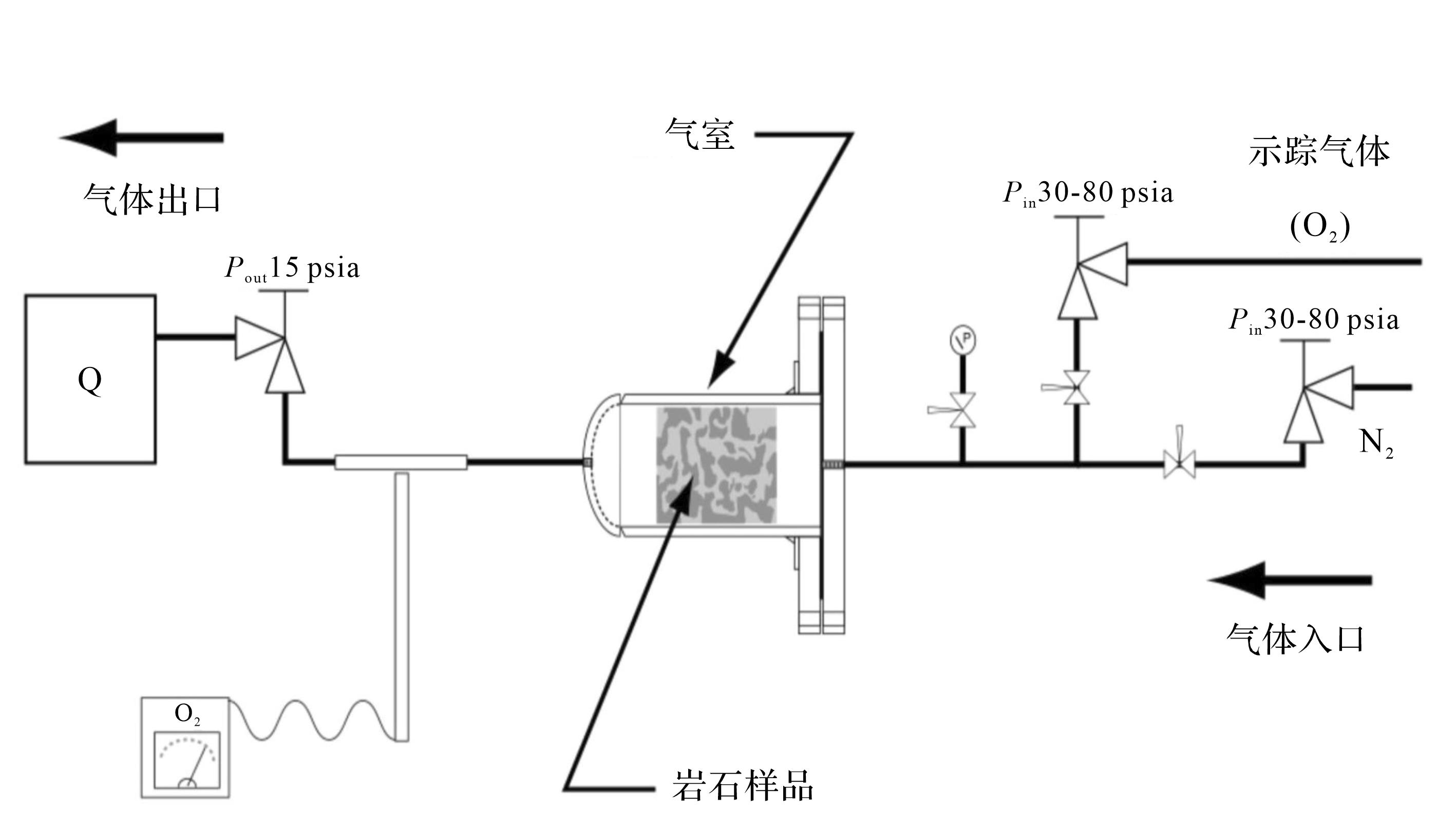
在物理模拟方面,加拿大学者Gingras et al.[57]做过一个实验,采用便携式探针渗透率仪(CoreLabs PP-250型),以氮气作为孔隙流体对流体传输特征进行测量。使用分散仪来测定体积渗透率和分散特性(图5)。在样品中建立N2气体的稳态流动,并在出口端测量气体体积。有效渗透率采用达西定律计算,该定律针对可压缩气体输运进行了修正。将一定浓度的O2作为示踪气体引入稳态流场,然后,使用手持式探测计随时测量出口处O2浓度(该仪器分辨率为千万分之一,精度为±1.5%)。这些数据被用来比较样本之间的相对分散程度[57,63]。

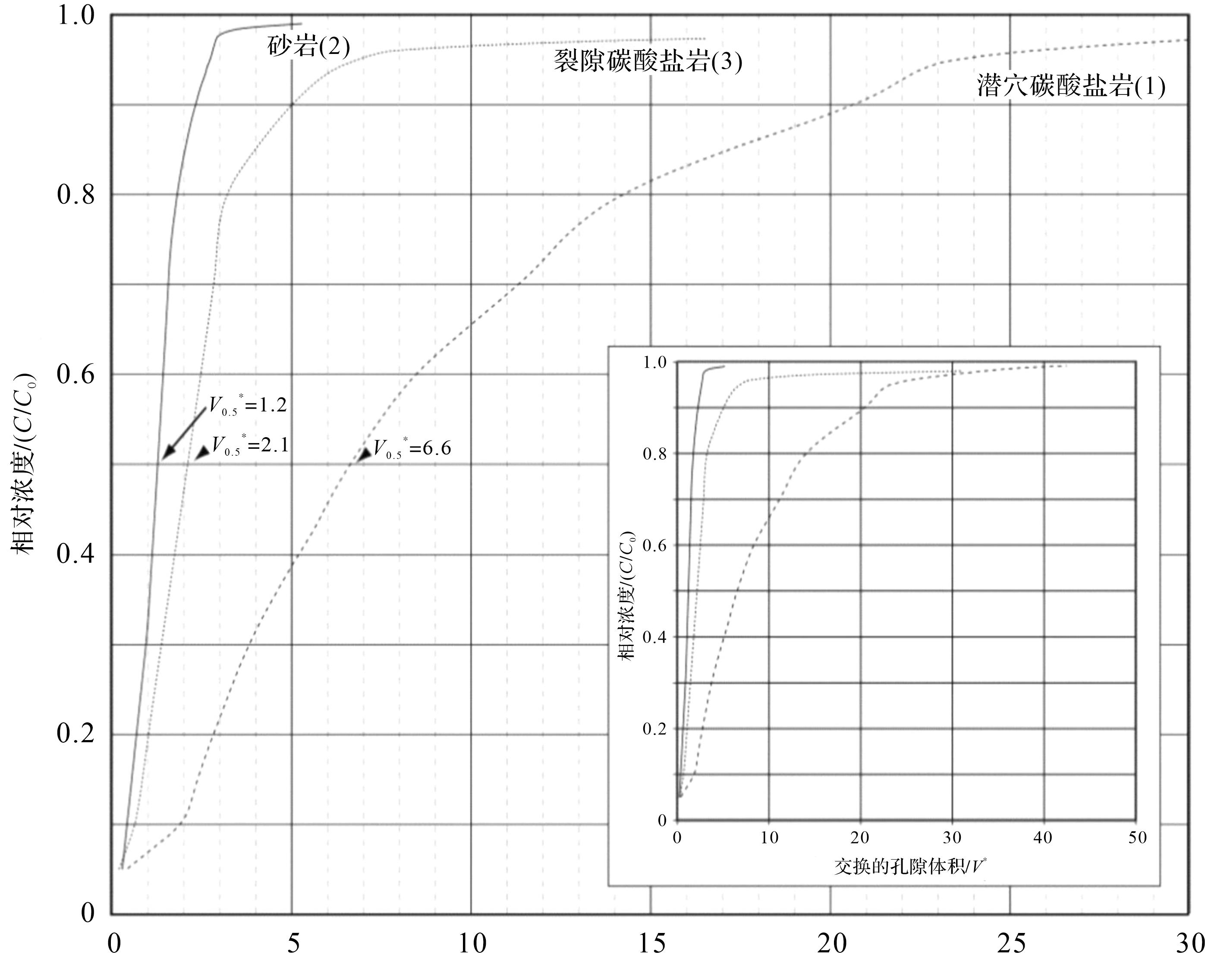
分散仪测量结果显示为示踪剂浓度(y轴)和充注气体的突破曲线(图6)。突破曲线显示了示踪气体(O2)浓度在出口处的增加速率。突破的发生是因为O2取代了最初用于充注孔隙的N2。突破曲线的形状与示踪剂波及的效率有关,也与岩石中渗透系统的性质有关。

图 6 分散仪流动实验结果:示踪气体相对浓度(C/C0)与交换的总孔隙体积(V* )附图显示了更大的孔隙体积交换范围,以更好地说明裂缝性和含生物潜穴碳酸盐岩所表现出的领先性(据文献[57]修改)
Figure 6. Results from the dispersometer flow experiments: relative tracer⁃gas concentration (C/C0) vs. gross pore volumes exchanged (V* ). The inset shows a broader range for pore volumes exchanged to better illustrate the leading exhibited by the fractured and the burrowed carbonate rocks (modified from reference [57])
该物理实验表明:生物扰动储层中流体的流动路径是弯曲的,流体在生物潜穴和围岩基质之间的交互作用要远远高于裂缝型碳酸盐岩和砂岩储层。识别这种流体路径尤为重要,因为这类储层的油气生产受储层介质的非均质性影响很大,特别是对油气二次回采方案的选取具有重要影响[56,64],这个模拟结果对于碳酸盐岩储层的开发具有重要意义,可以很好地说明目前生物潜穴储层采收率低的主要原因[65]。
除了物理模拟,目前还常用数值模拟研究生物扰动储层中流体的传输特征。数值模拟是对生物扰动地层的生物潜穴丰度、孔隙度和渗透率进行模拟[66],数值模拟可以分为宏观模拟传输特征和微观数值模拟流体的传输特征。在宏观数值模拟的流体传输特征实验方面,Eltom et al.[67]利用Petrel软件模拟Thalassinoides潜穴网络(Thalassinoides Burrow
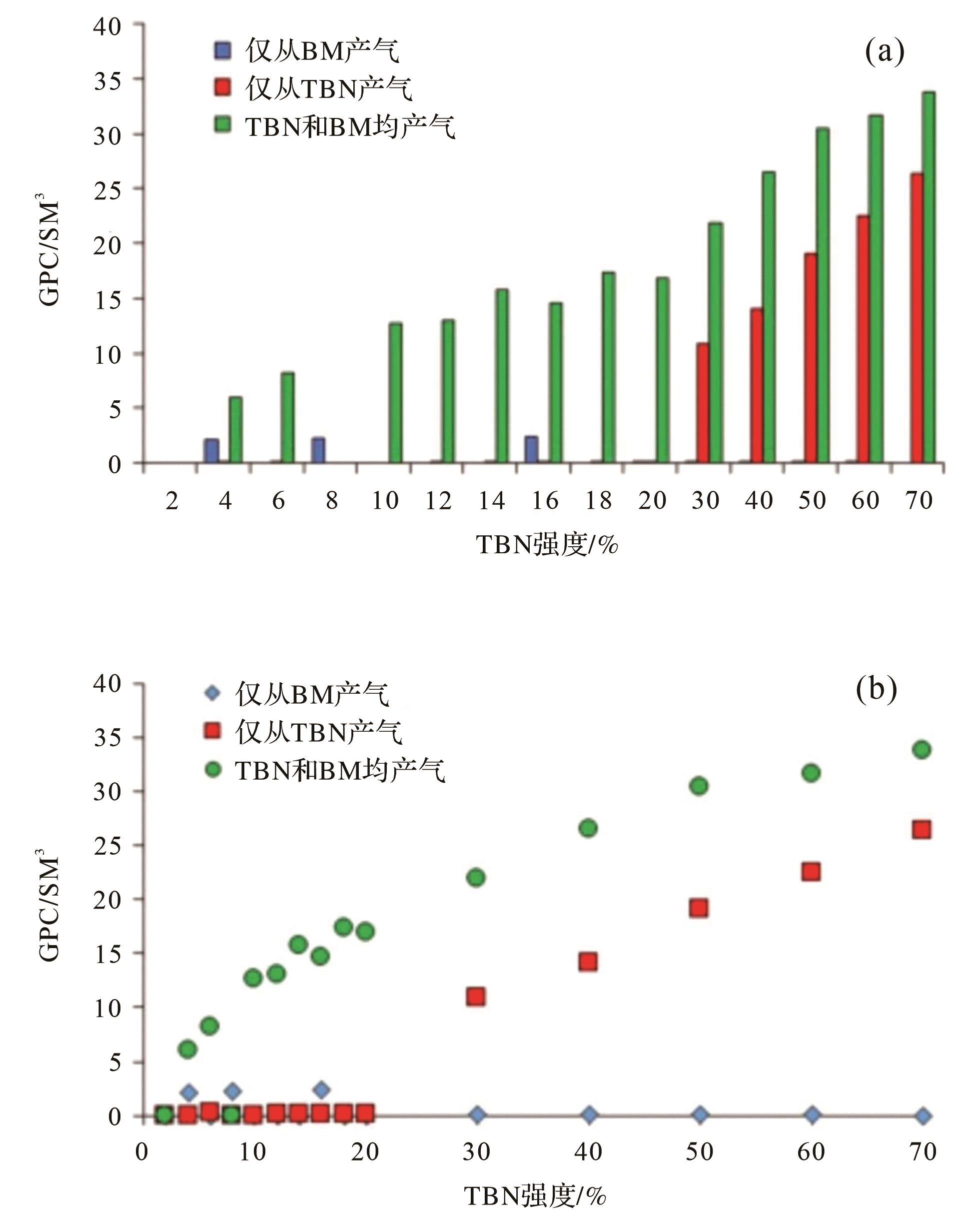
Networks,TBN)的流体传输特征,依据的是英国英格兰地区Birdsall露头上侏罗统的砂屑灰岩中的TBN。Eltom的数值实验模拟了三种情况:(1)关掉TBN的孔隙度(设置TBN的孔隙度设为0%),确保气流只能从潜穴的宿主围岩基质中产出;(2)关掉TBN宿主围岩基质的孔隙度(将潜穴基质的孔隙度设为0%),确保气体只能从TBN中产出;(3)将TBN和宿主围岩基质的孔隙度设置为一定的值,确保气体从TBN和潜穴基质中产出。 三种模拟情况下不同TBN强度(生物潜穴所占比例)累计产气的情况(图7),从a,b两幅图可以发现条件2和条件3,随着TBN强度(生物潜穴所占比例)的增加产气量都有较大的变化,确保气流只能从生物潜穴的宿主围岩基质中产出的模拟情况下,累计产气量变化不大。并且在气体仅从TBN内产出下,TBN的强度达到30%的时候才出现具有工业开采价值的产气量(>10 SM³),而在气体从TBN和基质中都产出情况下,TBN的生物扰动强度到10%的时候,就可以达到具有工业开采价值的产气量(>10 SM³)[67]。

三维TBN模型表明:潜穴的连通性取决于生物潜穴占比[68],一个完善的、连通的生物潜穴网络可以在只有12%的生物潜穴占比下建立起来。TBN的流体流速主要由TBN连通性和基质渗透率控制。宿主围岩基质与TBN之间的相互作用似乎连接了更多孤立的TBN体积,可能有助于气体从围岩基质扩散到TBN[67]。这些连通性模式决定了生物潜穴相关储层的渗透率是各向同性还是各向异性[68]。数值模拟为研究生物扰动储层中流体的传输特征提供富有成效的方法[69⁃71]。
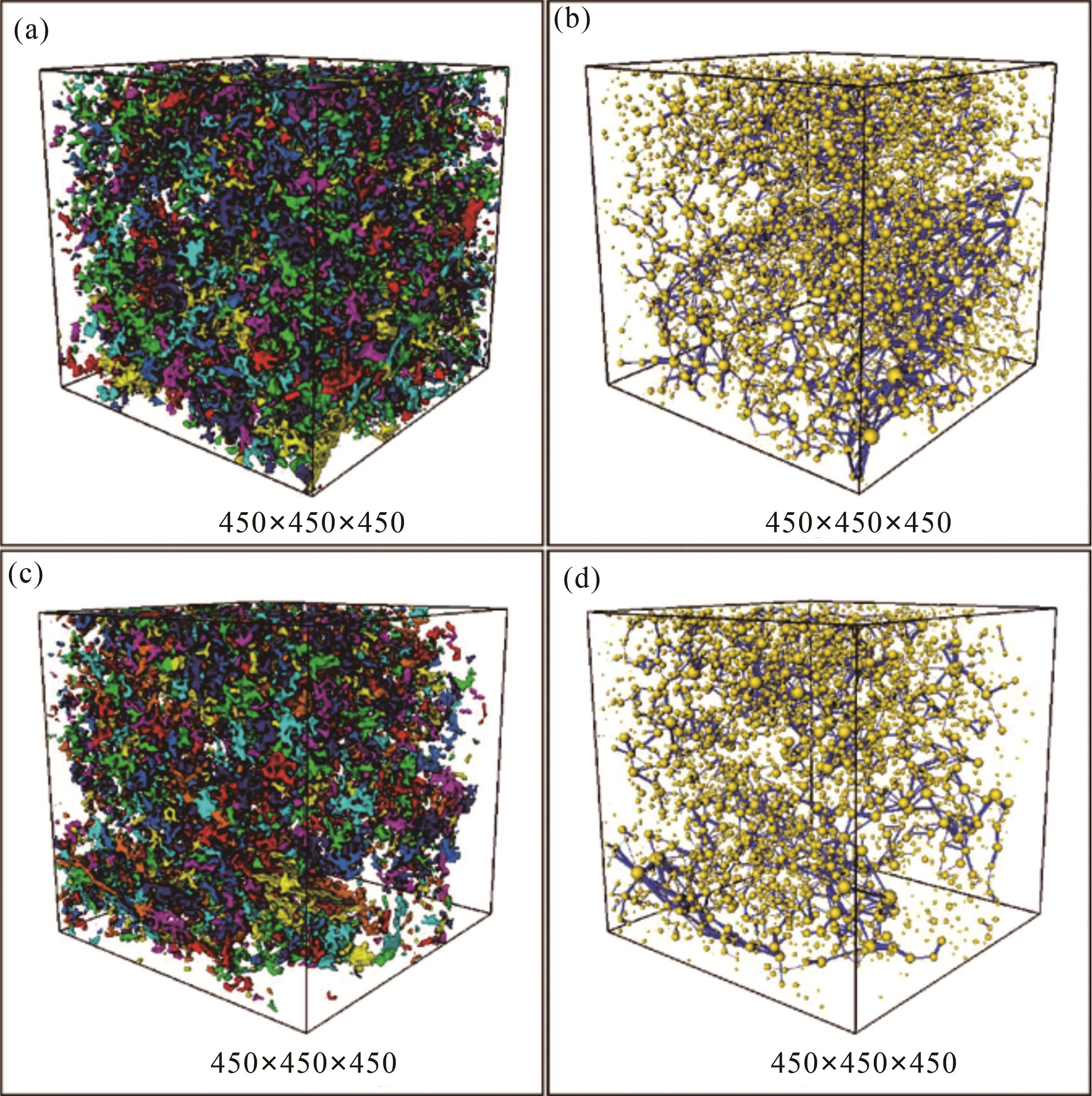
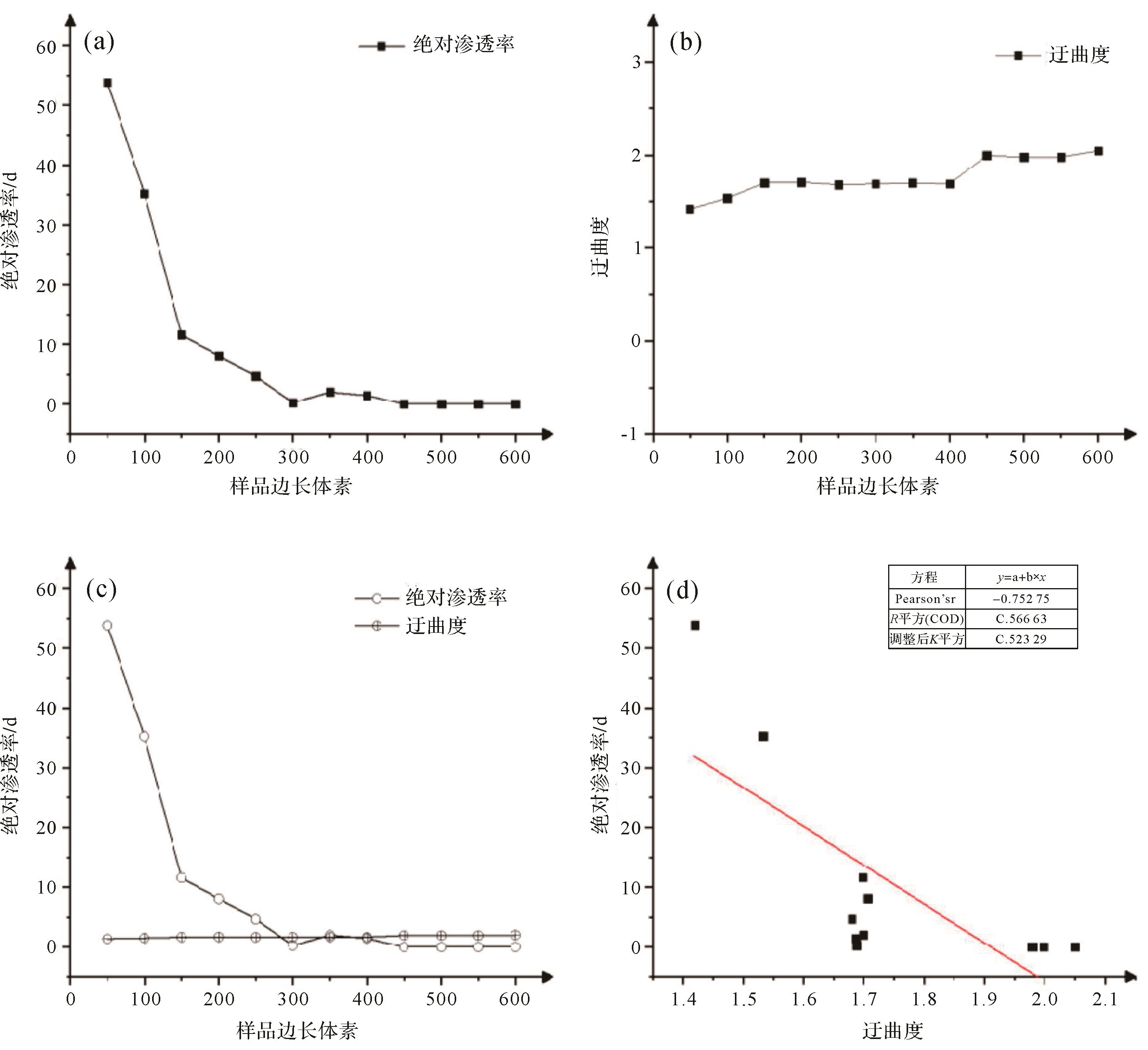
在微观数值模拟的流体传输特征实验方面,牛永斌等[19]选取塔河油田奥陶系生物扰动储层样品,使用X射线对岩石进行扫描成像[19,67,72],利用数字岩心软件三维重构出生物扰动储层的微观孔隙结模型,再进行生物扰动储层的微观渗流数值模拟(图8,9)。微观渗流数值模拟发现,随着样品尺寸的增大,样品孔隙度降低,渗透率也随之降低,这是因为岩心样品尺寸越大,潜穴的围岩基质就越多,这些围岩基质没有渗透性。样品尺寸越大,其孔隙度与渗透率越低,因为受到了围岩基质的影响。通过对渗流模拟得到的迂曲度与绝对渗透率的数据进行的分析能看出,渗透率与样品尺寸为负相关关系,迂曲度与样品尺寸呈正相关关系,从迂曲度和渗透率两者相关性分析发现,样品模型的迂曲度和渗透率是具有一定的相关性的。孔隙迂曲度和渗透率呈反比关系且相关性较强(图10)。

图 8 塔河油田奥陶系典型岩心样品孔隙结构定量表征工作流程
Figure 8. Workflow for the quantitative characterization of the pore structure of typical Ordovician core samples from Tahe oilfield

图 9 塔河油田奥陶系典型岩心样品孔隙空间网络模型的提取
Figure 9. Pore space network model of typical Ordovician core samples in Tahe oilfield

图 10 塔河油田奥陶系典型岩心样品迂曲度与绝对渗透率的数据
Figure 10. Tortuosity and absolute permeability of typical Ordovician core samples in Tahe oilfield
生物扰动的影响也在核磁共振岩心柱塞尺度上被识别出来。以大孔喉类为主导的两种不同孔喉类的双峰特征行为表明:生物潜穴提升了储层的孔喉连通性[12]。前人研究表明许多因素直接影响生物扰动油藏的体积流量参数。这些特征包括生物潜穴和基质之间的渗透性比值、生物扰动强度、潜穴连通性、潜穴表面积和潜穴结构,将整体渗透率估计与生物扰动强度联系起来时,应考虑生物潜穴连通性[73]。要进行全面综合的研究(包括沉积学、开发技术、岩石物理分析和地质统计建模)来了解储层质量[61]。
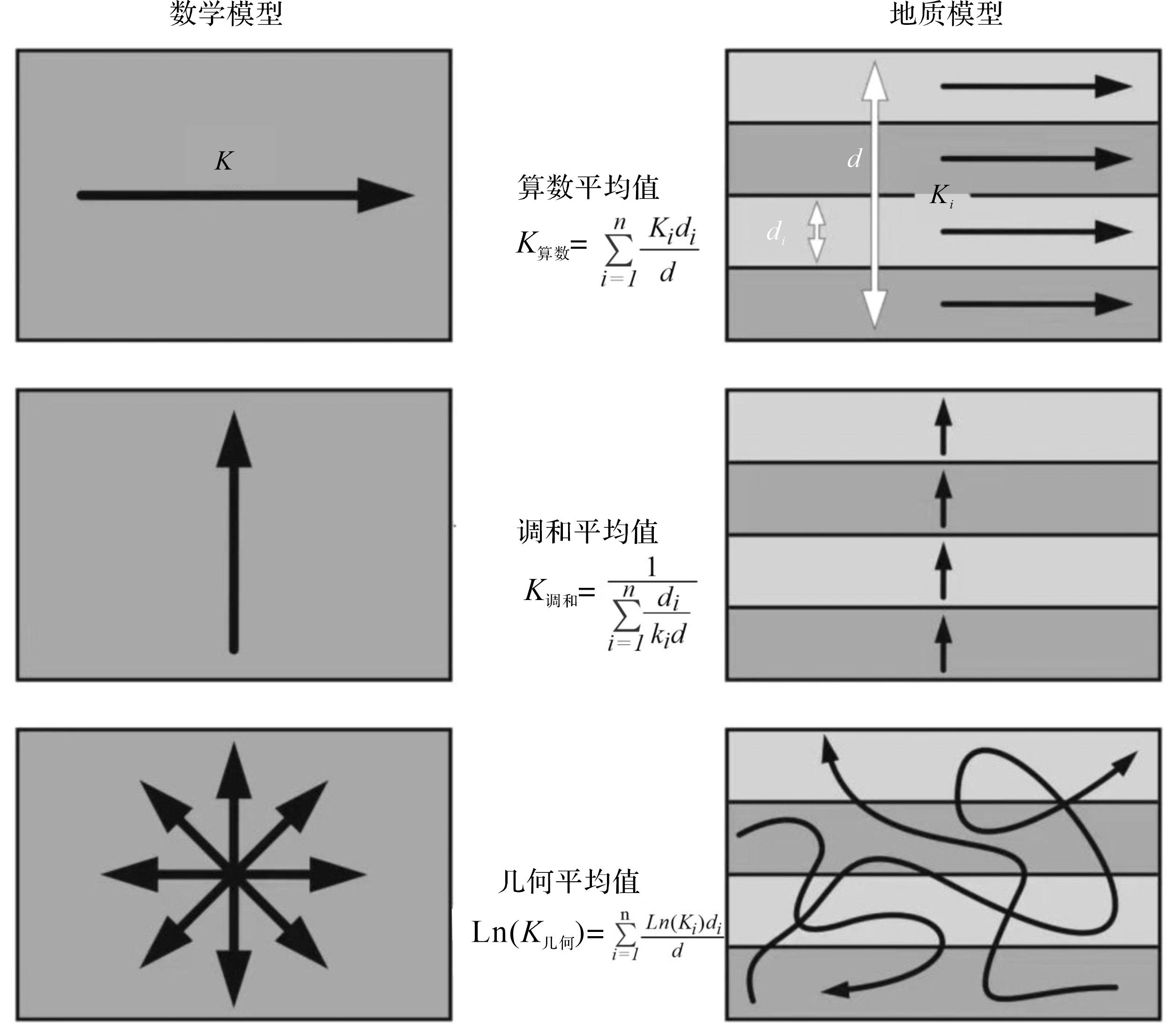
在计算地层渗透率中,有三个计算公式可以使用。它们分别适用于三种生物扰动储层介质[74](图11)。在潜穴低连通性的情况下,整体渗透率可以使用调和平均值来表征。然而,随着潜穴网络连通性越来越好,需要应用一种改进的算术平均值。在潜穴网络过渡性情况下,流体可渗透的生物潜穴结构只是零星连接,则广泛采用几何平均值,加权体积(di /d)以潜穴所占体积表示,与生物潜穴占比成正比[74]。

(1) 在平行于地层总渗透率,高潜穴连通性的条件下[75⁃76],确定沿连续平行渗透率域的平均体渗透率(K)由算术平均值(式(1))表示[56,77⁃78]:
(1) 式中:Ki 为岩性渗透率,d为地层厚度,di /d为加权体积(即各岩性所占体积的比例)。
(2) 在低潜穴连通性的条件下,垂直地层的总渗透率(K),由调和平均值(式(2))表示[75,79]:
(2) (3) 用于过渡性情况,即可渗透的潜穴结构仅局部相连的非层状地层的总渗透率(K),由体积加权几何平均值(式(3))表示[75,80]:
(3) -
对生物扰动储层进行分类及其属性研究,对储层预测和储层表征具有重要意义。Gingras et al.[55]根据沉积学、遗迹学、成岩作用和地层背景将生物扰动储层介质分为五大类:(1)界面控制的生物扰动储层(SCD);(2)非界面控制的生物扰动储层(NCD);(3)边界弱清晰的生物扰动储层(WDH);(4)隐蔽性生物扰动储层(CBH);(5)成岩成因的生物扰动储层(DTH)[54⁃56](图12)。
-
界面控制的生物扰动储层在固底和硬底暴露表面上很常见。这类生物扰动储层的最常见特征包括无衬里的潜穴,从单一表面下降到坚固的沉积物(压实基质),生物潜穴包含与围岩基质岩性不同的物质[81⁃82],并且通常不会在整个界面上普遍存在[83⁃84]。由于充填砂的生物潜穴位于泥岩基质中(或充填白云石的生物潜穴位于灰岩基质中),界面控制的生物扰动储层介质通常为双渗透率生物成因储层介质[56,74]。
界面控制的生物扰动储层在碎屑岩中常见,碳酸盐岩极少。这类储层受控于沉积不连续面(海侵面/潮到侵蚀面),一般生物潜穴发育在低渗透基质中后被粗粒沉积物充填,比如泥岩中的生物潜穴被砂质充填;生物潜穴类型一般与Glossifugites遗迹相关系密切,常见Skolithos,Diplocraterion等生物潜穴,常与Thalassinoides和Zoophycos交切。此外,深海浊积扇短期置居窗内广泛发育的生物潜穴也属于这一类。这类生物扰动储层一般为双渗透率生物成因储层介质;储层渗透率大小受控于潜穴渗透率与围岩基质渗透率的比值、潜穴连通性和潜穴密度。一般生物潜穴直径为0~10 cm,长度为0~70 cm,极端情况下可达3 m,平面延伸100 m至数千米。这类生物扰动储层的沉积环境有泥质潮坪、泥质滨岸和浅海陆棚等[56,74]。
Gingras et al.[56]对美国华盛顿州Willapa湾更新统的岩石进行测试,确定了三个变量控制Glossifungites遗迹相界面的流动特征:(1)潜穴渗透性(Kb )和基质渗透性(Km )之间的对比程度;(2)潜穴互渗量;(3)潜穴密度。计算机模拟结果表明,主要影响因素是渗透率对比(Kb /Km )和生物潜穴连通性[76,85]。这类生物扰动储层的典型案例还有:沙特阿拉伯Ghawar油气田侏罗系Hanifa组和加拿大英属哥伦比亚省三叠系Baldonnel组[56,74]。
-
非界面控制的生物扰动储层在碎屑岩常见,碳酸盐岩也有。这类生物扰动储层不受沉积界面控制,生物潜穴在低渗透沉积基质中形成后被粗粒沉积物充填(如泥岩基质中的生物潜穴被砂质沉积物充填),一般受风暴、浊流沉积影响;常与Cruziana遗迹相和Zoophycos遗迹相关系密切,有时在Nereites遗迹相中也可见到。常见的生物潜穴类型有Thalassinoides,Planolites,Chondrites,Palaeophycus和Zoophycos等。这类储层一般呈现为双渗透率生物成因储层介质;储层渗透率受控于生物潜穴与围岩基质的比值、潜穴连通性和潜穴密度。生物扰动程度可非常高,一般潜穴直径为0~5 cm,长度不等,厚10~100 m;平面延伸1~10 km。这类储层的形成沉积环境有浅海与浅海过渡带、潟湖、海湾、碳酸盐岩台地等[56,74]。
非界面控制的生物扰动储层的离散非均质性由被包裹在细粒基质中的生物潜穴组成,但并不局限于单个地层界面。这些生物潜穴结构在横向和纵向上是连续的,在近海、潟湖和海湾沉积环境中都很常见。由于生物潜穴与基质的岩性差异较大,通常存在双孔隙度与双渗透率混合生物成因储层介质。在近海和潟湖环境中,生物扰动强度可能相当高,潜穴占沉积物体积的30%~90%。垂直和水平潜穴的混合结构在Cruziana和Zoophycos遗迹相中典型发育。因此,尽管生物潜穴和基质之间的岩性差异很大,这类储层介质却可以变得非常各向同性[76]。高渗透区域受到生物扰动的位置和性质的影响,生物扰动改变了原始沉积物。
印度尼西亚东Java海域Sirasun-Sirasum气田新近系Paciran组发育这类生物扰动储层[86]。Paciran组分为下部砂岩段和上部有孔虫石灰岩段。Noble et al.[87]的研究表明,Paciran灰岩段是一个深海中上层碳酸盐岩矿床,由伊利石—蒙脱石黏土基质和球状有孔虫试验组成。研究还发现石灰岩段的垂直渗透性高于水平渗透性,证实了Zoophycos生物潜穴网络具有更高的渗透性[54]。这类生物扰动储层的典型案例还有:挪威海域白垩系Lysing组、东爪哇海新近系的Paciran组和中国南海琼东南盆地的三亚组储层等[22,56,74]。
-
边界弱清晰的生物扰动型储层在碎屑岩中常见,碳酸盐岩中少有。这类生物扰动储层是砂质基质发育生物潜穴后被粗粒沉积物充填(如砂岩基质中的生物潜穴被更粗砂质充填,生物潜穴充填物质分选性一般比基质要好);常见生物潜穴类型有Planolites,Thalassinoides,Ophiomorpha,Skolithos,Phycosiphon和一些Macaronichnus等。这类生物扰动储层一般呈现双孔隙度生物成因储层介质;储层渗透率与基质粒度、潜穴连通性和潜穴密度有关,生物扰动程度可非常高。一般潜穴直径为0~10 cm,长度不等,厚度达10 m;平面延伸100 m至数千米。这类生物扰动储层的沉积环境有前滨下部、临滨上部、三角洲和砂质海湾[56,74],生物潜穴充填物的分选性和堆积方式一般比围岩基质的要好。然而,由于岩性的相似性,潜穴和基质之间的渗透率差异通常小于两个数量级,因此属于典型的双孔隙度生物成因储层介质[74]。
加拿大萨斯喀彻温省西南部的白垩系Hatton气田的Alderson段,主要由泥质单元内连续的、横向广泛的薄层细粒砂岩组成,沉积环境为三角洲[88]。该储层单元发育许多砂质充填的生物潜穴。Lemiski et al.[88]使用点渗透率分析仪对该储层进行了测试分析,结果表明该段生物扰动储层为典型的双孔隙度生物成因储层介质,生物扰动对Alderson段天然气总体储量和产量的贡献巨大。这类生物扰动储层的典型案例还有:美国阿拉斯加州三叠系的Sag River组、挪威西部外海侏罗系的Ula组、加拿大纽芬兰岛白垩系的Ben Nevis组、Albert省白垩系的Niobrara组Medicine Hat段、中国塔里木盆地泥盆系的东河塘砂岩段、中国南海珠江口盆地的珠江组等[20,56,74]。
-
隐蔽性的生物扰动储层主要发育在碎屑岩中(砂岩中最常见)。一般为较小动物群扰动沉积层(残余和全部);最常见的生物就是Macaronichnus生物潜穴的造迹生物[7,89⁃90];因此,最常见的生物潜穴为Macaronichnus等。这类储层一般为双孔隙度生物成因储层介质;储层渗透率与围岩基质、潜穴充填物粒度相关,生物扰动程度极高。储层厚10~100 m;平面延伸100 m至数千米。这类生物扰动储层的形成沉积环境有障壁岛、三角洲内分流河道和临滨上部砂体等[56,74]。
隐蔽性生物扰动产生的岩石组构可能对储层的渗透率和孔隙度产生重大影响[74,91⁃93]。在一些生物扰动储层系统中,既存在隐蔽性的生物扰动砂岩,也存在未被生物扰动的砂岩,微型渗透率分析仪测试结果研究表明:未被生物扰动砂岩中保留了较强非均质性,导致了更广泛的渗透率值区间。相比之下,隐蔽性的生物扰动砂岩段非均质性就不那么明显。隐蔽性生物扰动作用可能不会导致渗透率增强,但会降低储层内部的非均质性,导致渗透率值分布更加均匀[74,93]。
Gordon et al.[92]在研究加拿大Albert省下白垩统Bluesky组储层时发现,该储层受到了Macaronichnusseggregatus的强烈生物扰动。薄片和扫描电子显微镜(SEM)分析显示,Macaronichnus seatus的造迹生物建造潜穴时避开了富含铁的岩石碎屑,在这个过程中导致了砂质沉积物的重新排序,生物潜穴充填物中含有较高的硅质成分(主要为石英),并且这些硅质颗粒边缘没有形成封堵孔隙的石英次生加大边,因此孔隙度和渗透率相对于宿主围岩基质较高(Macaronichnus发育区现场渗透率测量值比相似粒度的层状砂岩高出4倍),该生物扰动储层为双孔隙度生物成因储层介质。隐蔽性的生物扰动储层的典型案例还有:挪威西部外海侏罗系的Ula组、英国北海油田Bruce区块中侏罗统Tabert组储层和埃及法雍地区的上始新统储层等[56,74,94]。
-
成岩成因的生物扰动储层主要发育在碳酸盐岩中(尤其在白云质斑状灰岩中发育),在碎屑岩中比较少(仅在结核状砂岩中有该类储层的记录)。这类储层的形成主要与生物潜穴诱导的白云化作用、溶蚀作用和胶结作用有关;常见遗迹化石有Thalassinoides,Ophiomorpha,Balanoglossites,Chondrites等。一般为双渗透率或双孔隙度生物成因储层介质;该类储层渗透率与成岩特征、潜穴连通性和潜穴密度有关,生物扰动程度非常高。储层内一般潜穴直径为0~30 cm,长度不等,厚10~100 m;平面延伸1 km至数千米。该类储层形成的沉积环境主要为碳酸盐岩台或碳酸盐岩斜坡[56,74]。
成岩作用对生物潜穴相关碳酸盐岩层序的影响通常是极端的,可能包括许多不同的矿物(如菱铁矿、白云石、方解石、黄铁矿等)[74]。成岩蚀变生物潜穴的共同特征包括:(1)生物潜穴壁被胶结;(2)生物潜穴充填物的沉淀或溶解;(3)原始生物潜穴周围存在成岩晕;(4)潜穴内充填物的固结[3,36,95⁃96]。在某些情况下,生物潜穴充填物的矿化作用可以经历多个阶段,直至形成普遍胶结[58]。考虑到生物潜穴和围岩基质之间存在成岩作用的极大差异,该类生物扰动碳酸盐岩储层很容易在很短的横向和垂直距离内从双孔隙度生物成因储层介质变为双渗透率生物成因储层介质[97]。
Baniak et al.[17]在对加拿大Albert省上泥盆统Wabamun群的研究表明:白云化生物潜穴与非白云质灰泥—碎屑岩之间的边界非常清晰,并常常以低振幅的缝合线分界。生物潜穴充填物是以结晶程度较高的自形、半自形白云石晶体为主,它形白云石较为少见,晶体之间普遍可见基质灰岩残留;围岩基质则多为微晶方解石;这些白云化的潜穴充填物相对于周围的灰岩基质具有明显更高的渗透率[53]。目前,有一些学者试图用数值实验模拟来探索这类生物扰动储层渗透率分布和流体的运移特征[10,79]。成岩成因的生物扰动储层的典型例子还有:加拿大Manitoba省的Red River组[98]、Saskatchewan省Tyndall灰岩中的Selkirk段[99⁃100]、加拿大Williston盆地内奥陶系的Yeoman组、加拿大西北部泥盆系的Lonely Bay组、Albert省泥盆系的Palliser组以及中国塔河油田奥陶系鹰山组生物扰动储层等[18⁃19,56,74]。
-
近年来,随着新技术和新方法正在遗迹学研究中得到广泛应用,必将推动遗迹学在油气水储层领域应用的快速发展。如刘航宇等[101]通过自然伽马测井(GR)、侵入带电阻率测井(RHOB)、声波测井(DT)、补偿中子测井(NPHI)、侧向电阻率测井(RILD)和微球聚焦电阻率测井(MSFL)联合识别了生物扰动高渗透带(HPZ),并建立了基于RHOBnor/DTnor、RILD*MSFL和GR三个参数的测井识别模型。此外,通过对生物扰动高渗透带(HPZ)的岩性、孔隙类型、岩石物性、孔隙结构、层序地层学和早期成岩特征的研究,还揭示了生物扰动高渗透带(HPZ)的综合特征和地质演化过程。因此,将多源测井数据应用于生物扰动储层研究,识别出生物扰动储层的空间展布,能够更精确地确定整个油田的横向范围和垂向厚度。另外,将生物扰动储层的多源测录井资料与数字建模相联系,建立评价潜在储集岩质量、模拟盆地和含油气系统的地质模型,可以改善产量预测和提高采收率作业;而对生物扰动油气储层的多物理场、多化学场、多尺度和多相态的渗流模拟,可更好地理解油气水等流体在多孔储层介质中的渗流机理,这可为油田制定完井/穿孔决策和开发方案优选等方面提供高质量的指导。
值得说明的是,将能够表征大范围遗迹学信息的遗迹相和表征精细遗迹学信息的遗迹组构进行综合分析,比进行单方面的分析更能够产生一个好的、可信的结果;而将遗迹相和沉积相研究相结合,可提高生物扰动储层的空间分布预测的准确性。众多研究表明,将多种物理成因、化学成因和生物成因的沉积构造相联系是精确重建(古)环境的有效方法[52,94,102⁃105];通过识别岩石记录中的底栖生物行为可大大提升环境解译的准确性;而将生物扰动的演变特征、生物扰动模式和古生态参数的记录相结合,可以深刻揭示(古)生物/沉积的交互作用与后期的成岩改造过程,这可为沉积环境和成岩机理分析提供重要的相标志和生物学信息。上述多方面的综合研究毫无疑问对丰富储层地质学理论、实现油气增储、上产和提高采收率等方面具有重要意义。
-
(1) 生物扰动完全可以形成有效的油气水储层。生物扰动碎屑岩储集层中,典型的储集空间主要为原生粒间孔、次生溶蚀孔和生物铸模孔等,早期的原生孔隙因为经历了广泛的成岩蚀变被方解石胶结,差异型充填方解石胶结物显著降低了孔隙度和渗透率,但在随后的成岩过程中方解石胶结物又被溶解,使得粒间孔隙空间得以保留。生物扰动碳酸岩岩储集层中,典型的储集空间主要为原生粒间孔、晶间孔和溶蚀孔等,同生期—早成岩期溶蚀作用是生物扰动碳酸盐岩储集空间发育的关键因素,后期的白云化作用也对改善生物扰动碳酸盐岩储集性能具有积极意义。
(2) 根据生物扰动储层中流体的传输特征,生物扰动形成的岩石组构可以划分为两类储层介质:双孔隙度生物成因储层介质和双渗透率生物成因储层介质。生物潜穴充填物和基质渗透率比、生物扰动强度和连通性、潜穴表面积、潜穴的构型(结构与空间形态)特征是影响生物扰动储层储集性能的主要因素。目前,关于生物扰动储层中流体的传输特征研究主要有物理实验模拟和数值实验模拟两种研究思路。
(3) 根据沉积学、遗迹学、成岩作用和地层背景将生物扰动储层介质分为五大类:界面控制的生物扰动储层(SCD),非界面控制的生物扰动储层(NCD),边界弱清晰的生物扰动储层(WDH),隐蔽性的生物扰动储层(CBH),成岩成因的生物扰动储层(DTH)。各类生物扰动储层中生物潜穴充填物和基质渗透率比、生物扰动强度、潜穴连通性、潜穴表面积、潜穴的构型特征是影响储层物性的主要因素。
(4) 目前,尽管生物扰动储层的共同特征已被充分记录,但国内外对于生物扰动油气水储层的研究还处于初级阶段。可以预见的是,将沉积学、古生物地层学、地球物理学、储层地质学和地质建模结合起来,对生物扰动地层的生物潜穴丰度、孔隙度和渗透率等进行物理和数值实验模拟,可以帮助确定影响生物扰动油气水储层渗透率和产能的关键参数和变量,这对丰富储层地质学理论,实现油气增储、上产和提高采收率具有重要意义。
A Review and Perspective of Bioturbated Hydrocarbon and Water Reservoirs
-
摘要: 目的 在油气水储层中,生物扰动通过改变原生沉积组构降低或提高地层的孔隙度和渗透率,进而影响储层物性和流体流动特征。 方法 为了调研生物扰动储层的研究现状及进展,结合前人和笔者团队近年的研究成果,系统梳理和归纳总结了生物扰动油气水储层的储集空间类型、孔隙结构特征、储层介质类型、流体传输特征、储层类型及其属性特征。 结果 (1)生物扰动储层的储集空间类型包括:原生粒间孔、粒内溶孔、粒间溶孔和晶间孔等;(2)生物扰动储层可划分为双孔隙度生物成因储层介质和双渗透率生物成因储层介质,两类储层介质在生物潜穴类型、运移动力、流体传输特征、沉积环境等方面存在不同程度的差异;(3)生物扰动储层包括界面控制的生物扰动储层、非界面控制的生物扰动储层、边界不清晰的生物扰动储层、隐蔽性生物扰动储层、成岩成因的生物扰动储层五种类型,潜穴充填物和基质渗透率比、生物扰动强度和连通性、生物潜穴表面积、潜穴的构型特征是影响生物扰动储层岩石物性的主控因素。 结论 尽管生物扰动储层的共同特征已被充分记录,但国内外对于生物扰动储层的研究还处于初级阶段,但可预见的是加强生物扰动储层的研究对丰富储层地质学理论,实现油气增储、上产和提高采收率具有重要意义。Abstract: Objective In bioturbated hydrocarbon and water reservoirs, bioturbation changes the petrophysical properties of the host rocks by changing the primary sedimentary fabric, reducing or increasing the porosity and permeability of the primary formation, which in turn affects the reservoir petrophysical properties and fluid flow characteristics. To investigate the research status and progress of bioturbated hydrocarbon and water reservoirs, Methods combined with recent research results, the reservoir space types, pore structure characteristics, reservoir media nature, fluid transport characteristics, reservoir types, and their property characteristics of bioturbation hydrocarbon and water reservoirs have been sorted and summarized. Results The results show that: (1) The reservoir space types of bioturbated reservoirs include: primary intergranular, intra-grain dissolved, inter-grain dissolved, and intergranular pores. In bioturbated clastic rock reservoirs, typical reservoir spaces are primary intergranular, secondary dissolution, and biological mold pores. The early primary pores were filled with calcareous cements to reduce porosity and permeability, but in the subsequent diagenesis, if these calcareous cements were dissolved, these intergranular pores could be preserved. In bioturbated carbonate reservoirs, typical respace spaces contain primary intergranular, intercrystalline, and dissolution pores. Syngeneic or early diagenetic dissolution is the key factor in the development of bioturbated carbonate reservoir space. (2) Based on the relationship between the permeability of burrows and the host sediments, the bioturbated reservoir can be divided into dual-porosity and dual-permeability biogenic reservoir media, and the two types of reservoir media have significant differences in burrow types, migration dynamics, fluid transport characteristics, and depositional environment. At present, there are two main research approaches to the study of fluid transport characteristics in bioturbation reservoirs: physical and numerical simulation. The physical simulation shows a broader range for pore volumes exchanged to better illustrate the leading exhibited by burrowed carbonate rocks. The numerical simulation reveals a well-connected burrow network can occur in strata of as little as 12% burrow intensity, and burrow connectivity increases nonlinearly with increasing burrow intensity. Numerical flow simulations of Thalassinoides Burrow Networks (TBN) models indicate that cumulative gas production (GPC) is controlled primarily by TBN connectivity and matrix permeability. With an impermeable matrix, production comes through TBN, starting only at a burrow intensity of 30%, and GPC increases linearly with burrow intensity. With a permeable matrix, and low permeability contrast between matrix and TBN (one order of magnitude) production starts at relatively low TBN intensity (4%), and GPC increases nonlinearly with burrow intensity. The interaction between matrix and TBN connects more isolated TBN bodies and increases GPC, likely helping the gas diffusion from matrix through TBN. The fluid flow simulation also illustrates the likelihood of the presence of permeability that drives early water breakthrough (super-K) at burrow intensity > 50%. The results advance the quantitative understanding of the impact of burrow connectivity on fluid flow properties of TBN-bearing strata and provide a workflow that can be used to model other burrow morphologies and understand their impact on flow properties of bioturbated reservoirs. (3) Based on the burrow characteristics of bioturbated reservoirs, fluid transport characteristics in reservoir media, and reservoir petrophysical characteristics, bioturbated reservoirs can be divided into five types: Surface-constrained, Non-surface-constrained, Weakly defined, Cryptic, and Diagenetic. The main controlling factors affecting the petroproperties of bioturbated reservoirs are analyzed in detail, including the permeability ratio of burrow-fills and hosting matrix, bioturbation intensity and connectivity, burrow-surface area, and burrow architecture. Conclusions Although the common characteristics of bioturbated reservoirs have been fully documented, the domestic and international relevant research on bioturbated hydrocarbon and water reservoirs is still in its primary stage; however, strengthening these studies is of great significance and imparts important implications for enriching the theory of reservoir geology, increasing oil and gas storage, and enhancing production and recovery.
-
Key words:
- bioturbation /
- hydrocarbon reservoir /
- reservoir space /
- porosity /
- permeability
-
图 1 Ben Nevis L⁃55岩心样品1,2,3切片和薄片详细信息(2 475~2 388 m)(据文献[28]修改)
(a)样品1的岩心切片(2 475 m),具有强烈的生物扰动(>90%)和蛇形洞穴;(b)过大的孔隙集中在Ophiomorpha生物潜穴的边缘;(c)大的蓝色超大孔隙的特写;(d)粒间孔隙率蓝色、被溶蚀的颗粒和颗粒状孔洞。样品孔隙率为20.3%;(e)样品2岩心切片(2 460 m),具有强烈的生物扰动(>90%)和Ophiomorpha生物潜穴;(f)生物潜穴斑点结构;(g)富含石英的极细砂岩;(h)粒间孔隙度(蓝色)和充填孔隙的黏土矿物;样品孔隙度为27.2%;(i)样品3岩心切片(2 388 m),具有强烈的生物扰动(>90%)、Ophiomorpha和Thalassinoides;(j)清洁的沙子,Thalassinoides 生物潜穴晕圈和泥质的Ophiomorpha生物潜穴衬壁;(k)Thalassinoides生物潜穴边缘特写;(l)粒间孔隙(蓝色)、黏土次生加大和充填孔隙的黏土矿物集中在生物潜穴衬壁中;样品孔隙度为23.4%;薄片(b~d、f~h和j~l)中注入了蓝色的环氧树脂,蓝色显示孔隙,灰白色为石英颗粒,深色为有机质、黏土矿物或黄铁矿;图a、图e和图i中的黑色虚线矩形表示获取薄片样本的位置,图b、c、f、g、j和k中的黑色虚线框表示拍摄单偏振光显微照片的位置
Figure 1. Porosity and petrographic details of samples 1, 2, and 3 from bioturbated sandstone from Ben Nevis L⁃55 (2 475⁃2 388 m) (modified from reference [28])
(a) core⁃slab sample 1 (2 475 m) with intense bioturbation (>90%) and Ophiomorpha burrows; (b) oversized pores are concentrated at Ophiomorpha burrow margins; (c) close⁃up of large gray over⁃sized pores; (d) interparticle porosity (gray), corroded grains, and grain⁃shaped voids. sample porosity is 20.3%; (e) core⁃slab sample 2 (2 460 m) with intense bioturbation (>90%) and Ophiomorpha burrows; (f) burrow⁃mottled texture; (g) quartz⁃rich very fine sandstone; (h) interparticle porosity (gray) and pore⁃filling clay minerals. Sample porosity is 27.2%; (i) core⁃slab sample 3 (2 388 m) with intense bioturbation (>90%), Ophiomorpha and Thalassinoides; (j) clean sand Thalassinoides burrow halo and mudstone⁃rich Ophiomorpha burrow lining; (k) close⁃up of Thalassinoides burrow margin; (l) interparticle porosity (gray), clay overgrowths, and pore⁃filling clay minerals concentrated in burrow lining. Sample porosity is 23.4%; Thin sections (b⁃d, f⁃h, and j⁃l) are impregnated with blue epoxy, gray shows pore space, off⁃white is quartz grains, and dark colors are organic matter, clay minerals, or pyrite; Dashed black rectangles in panels a, e, and i delineate where thin⁃section samples were taken, and dashed black boxes in panels b, c, f, g, j, and k delineate where plane⁃polarized light photomicrographs were taken
图 2 Ben Nevis L⁃55岩心样品4,5切片和薄片详细信息(2 357~2 356 m)(据文献[28]修改)
(a)样品4岩心切片(2 357 m),具有低生物扰动(5%~30%)、垂直定向的Ophiomorpha生物潜穴和纹层;(b)分散的纹层和模糊的生物潜穴;(c)黏土丰富的生物潜穴衬壁的特写;(d)颗粒间孔隙(灰色)和充填孔隙的黏土、方解石和黏土胶结物,封堵了可用的孔隙空间;样品孔隙度为4.9%;(e)样品5岩心切片(2 356 m),具有中等生物扰动(30%~60%)、Ophiomorpha生物潜穴、Chondrites生物潜穴和纹层;(f)泥岩充填的Chondrites在粉细砂岩中形成生物潜穴;(g)Chondrites特写;(h)Chondrites生物潜穴中的粒间孔隙度(蓝色)和充填孔隙的黏土矿物。样品孔隙度为16.8%;薄片(b~d、f~h)注入了蓝色的环氧树脂,蓝色为孔隙,灰白色为石英颗粒,深色为有机质、黏土矿物和黄铁矿。图a和图e中的黑色虚线矩形表示采集薄片样品的位置,图b、c、f和g中的黑色虚线框表示拍摄单偏振光显微照片的位置
Figure 2. Porosity and petrographic details of samples 4,5 from bioturbated sandstone from Ben Nevis L⁃55 (2 357⁃2 356 m) (modified from reference [28])
(a) core⁃slab sample 4 (2 357 m) with low bioturbation (5%⁃30%), vertically oriented Ophiomorpha burrow, and laminations; (b) diffuse laminations and indistinct burrow; (c) close⁃up of clay⁃rich burrow lining; (d) interparticle porosity (blue) and pore⁃filling clay, calcite, and clay cements occluding available pore space. Sample porosity is 4.9%; (e) core⁃slab sample 5 (2 356 m) with moderate bioturbation (30%⁃60%), Ophiomorpha burrow, Chondrites burrows, and laminae; (f) mudstone⁃filled Chondrites burrows in mud⁃rich, diffusely laminated very fine sandstone; (g) close⁃up of Chondrites; (h) Interparticle porosity (gray) and pore⁃filling clay minerals in Chondrites burrow; Sample porosity is 16.8%
图 3 双重孔隙度生物成因储层中的流体流动行为(据文献[56]修改)
(a)包裹在砂岩基质中的砂质充填的潜穴的岩心切片图像(Ophiomorpha和模糊的类Thalassinoides是不可见的);(b)带有生物扰动的沉积岩示意图显示为深灰色;(c)在双重孔隙流动介质中,流体通过基质和生物潜穴充填物进行平流;在生物潜穴渗透率较高的地方,存在更大的流体流动通量
Figure 3. Fluid⁃flow behavior in dual⁃porosity biogenic reservoir (modified from reference [56])
(a) slabbed core image of sand⁃filled burrows encased in a sandstone matrix (Ophiomorpha and vague Thalassinoides are discernible); (b) schematic sedimentary rock with bioturbation shown as darker gray; (c) within dual⁃porosity flow media, fluid is advected through both the matrix and the trace⁃fossil fill; Where the trace⁃fossil permeability is higher, a greater flux exists in the burrows
图 4 双渗透生物成因储层中的流体流动行为(据文献[56]修改)
(a)岩心切片图像显示石灰岩基质中的生物潜穴相关的白云石;(b)生物扰动沉积介质示意图显示为深灰色;(c)在双重渗透储层介质中,流体通过生物潜穴和基质进行平流;渗透率较低的基质仅通过扩散来促进流动
Figure 4. Fluid⁃flow behavior in dual⁃permeability biogenic reservoir (modified from reference [56])
(a) slabbed core image showing trace⁃fossil⁃associated dolomite in a limestone matrix; (b) schematic sedimentary medium with bioturbation shown as darker gray; (c) within dual⁃permeability flow media, fluid is advected through both the trace fossils and the matrix (dominantly by advection); The lower permeability matrix contributes to flow only via diffusion
图 6 分散仪流动实验结果:示踪气体相对浓度(C/C0)与交换的总孔隙体积(V* )附图显示了更大的孔隙体积交换范围,以更好地说明裂缝性和含生物潜穴碳酸盐岩所表现出的领先性(据文献[57]修改)
Figure 6. Results from the dispersometer flow experiments: relative tracer⁃gas concentration (C/C0) vs. gross pore volumes exchanged (V* ). The inset shows a broader range for pore volumes exchanged to better illustrate the leading exhibited by the fractured and the burrowed carbonate rocks (modified from reference [57])
Fig.6
图 7 三种模拟条件下的累积产气量(GPC)结果(据文献[67]修改)
实验条件1:仅从潜穴基质(BM)生产;实验条件2:仅从TBN生产;实验条件3:BM和TBN同时生产;(a)条形图,显示三种模拟条件下GPC之间的变异性;(b)三种实验模拟条件的产气量百分比绘制的GPC
Figure 7. Cumulative gas production (GPC) results for the three simulation conditions (modified from reference [67])
condition 1: production from burrow matrix (BM) alone; condition 2: production from TBN (Thalassinoides Burrow Networks) alone; condition 3: production from both BM and TBN; (a) bar chart showing variability among GPC of the three simulation conditions; (b) GPC plotted based on TBN percentages for each simulation condition
图 8 塔河油田奥陶系典型岩心样品孔隙结构定量表征工作流程
Figure 8. Workflow for the quantitative characterization of the pore structure of typical Ordovician core samples from Tahe oilfield
Fig.8
图 9 塔河油田奥陶系典型岩心样品孔隙空间网络模型的提取
(a)样品1孔隙分割模型;(b)样品1球棍模型;(c)样品2孔隙分割模型;(d)样品2球棍模型
Figure 9. Pore space network model of typical Ordovician core samples in Tahe oilfield
(a) sample 1 pore segmentation model; (b) sample 1 ball⁃and⁃stick model; (c) sample 2 pore segmentation model; (d) sample 2 ball⁃and⁃stick model
图 10 塔河油田奥陶系典型岩心样品迂曲度与绝对渗透率的数据
(a)样品边长体素与绝对渗透率交会图;(b)样品边长体素与迂曲度交会图;(c)样品边长体素与绝对渗透率、迂曲度交会图;(d)迂曲度与绝对渗透率交会图
Figure 10. Tortuosity and absolute permeability of typical Ordovician core samples in Tahe oilfield
(a) cross plot of sample length voxels and absolute permeability; (b) cross plot of sample length voxels and tortuosity; (c) cross plot of sample side length voxels with absolute permeability and tortuosity; (d) cross plot of tortuosity and absolute permeability

图 12 五类生物扰动储层特征及其分布,它们受强烈沉积环境的影响(据文献[74]修改)
界面控制的生物扰动储层不均匀沿坚硬底和硬底暴露面是常见的,图片来自美国华盛顿州Willapa湾的露头,代表了一个Glossifungities遗迹相例子;非界面控制的生物扰动储层在浅海和潟湖环境中很常见,图片来自挪威海上白垩统Lysing组的岩心,显示粉砂岩基质中充满砂岩的Thalassinoides和Planolites。边界不清晰的生物扰动储层在滨海下部和富砂的浅海沉积中很常见,图片来自挪威北海上侏罗统Ula组的岩心,显示了一组重复排列的垂直Ophiomorpha nodosa横切面,包含Ophiomorpha irregulaire 和Palaeophycus的斑状组构;隐蔽性的生物扰动储层在河道(河口湾和分支河道)和近岸岸线沉积(前滨、临滨上部、三角洲前缘)中很常见,图片来自加拿大艾伯塔省下白垩统Clearwater组的岩心,照片显示了一块隐蔽性生物扰动的斑状砂岩;成岩成因的生物扰动储层,从潟湖环境到近海环境均可发现,反映了同沉积到沉积后过程,图片来自加拿大萨斯喀彻温省一座政府大楼上露出的晚奥陶世Tyndall岩(红河组Selkirk段)照片,显示了白云化的生物洞穴
Figure 12. Characteristics and distribution of five types of bioturbated reservoirs, is highly influenced by the depositional environment (modified from reference [74])
Surface⁃constrained bioturbated reservoirs are common along firmground and hardground exposure surfaces; Outcrop photo from Willapa Bay, Washington, USA represents an example of a Glossifungities ichnofacies. Nonsurface⁃constrained bioturbated reservoirs are common in offshore and lagoonal settings; Core photo example from the Upper Cretaceous Lysing Formation, Norwegian Sea, showing sandstone⁃filled Thalassinoides and Planolites in a siltstone matrix. Weakly defined bioturbated reservoirs are common in lower shoreface and sand⁃rich proximal offshore deposits; Core photo example from the Upper Jurassic Ula Formation, Norwegian North Sea, showing an assemblage of heavily lined vertical Ophiomorpha nodosa cross⁃cutting a burrow mottled fabric containing Ophiomorpha irregulaire and Palaeophycus; Cryptic bioturbated reservoirs are common within channels (estuarine and distrib-utary) and paralic shoreline deposits (foreshore, upper shoreface, delta front); Core photo example from the Lower Cretaceous Clearwater Formation, Alberta, Canada showing a cryptic mottled sandstone; Diagenetic bioturbated reservoirs can be found from lagoonal to offshore settings and are a reflection of syn⁃to post⁃depositional processes; Photo example of an outcropping Late Ordovician Tyndall Stone (Selkirk member of the Red River Formation) from a government building in Saskatchewan, Canada showing dolomitized burrows
-
[1] Hülse D, Vervoort P, van de Velde S J, et al. Assessing the impact of bioturbation on sedimentary isotopic records through numerical models[J]. Earth-Science Reviews, 2022, 234: 104213. [2] Dey J, Sen S. Impact of bioturbation on reservoir quality and production: A review[J]. Journal of the Geological Society of India, 2017, 89(4): 460-470. [3] Niu Y B, Marshall J D, Song H B, et al. Ichnofabrics and their roles in the modification of petrophysical properties: A case study of the Ordovician Majiagou Formation, northwest Henan province, China[J]. Sedimentary Geology, 2020, 409: 105773. [4] Dorador J, Rodríguez-Tovar F J, Miguez-Salas O. The complex case of Macaronichnus trace fossil affecting rock porosity[J]. Scientific Reports, 2021, 11(1): 1975. [5] Taylor A, Goldring R, Gowland S. Analysis and application of ichnofabrics[J]. Earth-Science Reviews, 2003, 60(3/4): 227-259. [6] Moore D M. Impact of super permeability on completion and production strategies[C]//Proceedings of the SPE Middle East oil and gas show and conference. Bahrain: SPE, 1989: SPE-17974-MS. [7] 龚紫娟,张青田. 生物扰动影响沉积物理化特征的研究进展[J]. 海洋湖沼通报,2022,44(2):166-172. Gong Zijuan, Zhang Qingtian. Influence of macrobenthic bioturbation on the physic-chemical characteristics of sediment: A review[J]. Transactions of Oceanology and Limnology, 2022, 44(2): 166-172. [8] 杨伟芳,王波,张惠良. 塔里木盆地东河砂岩段中的生物扰动作用[J]. 新疆石油地质,2010,31(5):493-496. Yang Weifang, Wang Bo, Zhang Huiliang. Bioturbation of Donghe sandstone member in Tarim Basin[J]. Xinjiang Petroleum Geology, 2010, 31(5): 493-496. [9] 纪友亮,赵澂林,刘孟慧. 生物扰动构造对碎屑岩储层储集性能的影响[J]. 石油大学学报(自然科学版),1990,14(6):1-8. Ji Youliang, Zhao Chenglin, Liu Menghui. The effect of bioturbate structure on the petrophysical properties of sandstone reservoir[J]. Journal of the University of Petroleum, China, 1990, 14(6): 1-8. [10] Hsieh A I, Allen D M, MacEachern J A. Statistical modeling of biogenically enhanced permeability in tight reservoir rock[J]. Marine and Petroleum Geology, 2015, 65: 114-125. [11] Quaye J A, Jiang Z X, Zhou X W. Bioturbation influence on reservoir rock quality: A case study of well Bian-5 from the Second member Paleocene Funing Formation in the Jinhu Sag, Subei Basin, China[J]. Journal of Petroleum Science and Engineering, 2019, 172: 1165-1173. [12] Singh A, Jha N K, Mandal P P, et al. Pore throat characterization of bioturbated heterogeneous sandstone, Bhuj Formation, Kachchh India: An integrated analysis using NMR and HPMI studies[J]. Journal of Petroleum Science and Engineering, 2022, 211: 110221. [13] Loucks R G, Reed R M, Ruppel S C, et al. Morphology, genesis, and distribution of nanometer-scale pores in siliceous mudstones of the Mississippian Barnett Shale[J]. Journal of Sedimentary Research, 2009, 79(12): 848-861. [14] Chalmers G R, Bustin R M, Power I M. Characterization of gas shale pore systems by porosimetry, pycnometry, surface area, and field emission scanning electron microscopy/transmission electron microscopy image analyses: Examples from the Barnett, Woodford, Haynesville, Marcellus, and Doig units[J]. AAPG Bulletin, 2012, 96(6): 1099-1119. [15] Curtis M E, Cardott B J, Sondergeld C H, et al. Development of organic porosity in the Woodford Shale with increasing thermal maturity[J]. International Journal of Coal Geology, 2012, 103: 26-31. [16] Miguez-Salas O, Dorador J, Rodríguez-Tovar F J, et al. X-ray microtomography analysis to approach bioturbation's influence on minor-scale porosity distribution: A novel approach in contourite deposits[J]. Journal of Petroleum Science and Engineering, 2022, 208: 109251. [17] Baniak G M, Gingras M K, Pemberton S G. Reservoir characterization of burrow-associated dolomites in the Upper Devonian Wabamun Group, Pine Creek gas field, central Alberta, Canada[J]. Marine and Petroleum Geology, 2013, 48: 275-292. [18] 牛永斌,崔胜利,胡亚洲,等. 塔河油田奥陶系生物扰动型储集层的三维重构及启示意义[J]. 古地理学报,2018,20(4):691-702. Niu Yongbin, Cui Shengli, Hu Yazhou, et al. Three-dimensional reconstruction and their significance of bioturbation-type reservoirs of the Ordovician in Tahe oilfield[J]. Journal of Palaeogeography (Chinese Edition), 2018, 20(4): 691-702. [19] 牛永斌,徐资璐,刘圣鑫,等. 塔河油田奥陶系生物扰动碳酸盐岩储集层微观孔隙结构的数字化表征与连通性分析[J]. 古地理学报,2020,20(4):785-798. Niu Yongbin, Xu Zilu, Liu Shengxin, et al. Digital characterization and connectivity analysis of microcosmic pore structures of the Ordovician bioturbated carbonate rock reservoirs in Tahe oilfield[J]. Journal of Palaeogeography (Chinese Edition), 2020, 22(4): 785-798. [20] 牛永斌,崔胜利,胡亚洲,等. 塔里木盆地塔河油田奥陶系数字岩心图像中生物扰动的定量表征[J]. 古地理学报,2017,19(2):353-363. Niu Yongbin, Cui Shengli, Hu Yazhou, et al. Quantitative characterization of bioturbation based on digital image analysis of the Ordovician core from Tahe oilfield of Tarim Basin[J]. Journal of Palaeogeography, 2017, 19(2): 353-363. [21] de Araújo O M O, Aguilera O, Coletti G, et al. X-ray micro-computed tomography of burrow-related porosity and permeability in shallow-marine equatorial carbonates: A case study from the Miocene Pirabas Formation, Brazil[J]. Marine and Petroleum Geology, 2021, 127: 104966. [22] 牛永斌,程梦园,程怡高,等. 琼东南盆地北部新近系三亚组Ophiomorpha-Thalassinoides遗迹组构的储层改造效应[J]. 古地理学报, 2023,25(6):1407-1420. Niu Yongbin, Cheng Mengyuan, Cheng Yigao, et al. Reservoir modification effect of Ophiomorpha-Thalassinoides ichnofabric in the Neogene Sanya Formation in northern Qiongdongnan Basin[J]. Journal of Palaeogeography, 2023, 25(6): 1407-1420. [23] 赵佳如,牛永斌,王敏,等. 塔河油田奥陶系生物扰动型碳酸盐岩储集层特征及其孔隙度计算样本检验模型[J]. 沉积学报,2021,39(2):482-492. Zhao Jiaru, Niu Yongbin, Wang Min, et al. Reservoir characteristics and porosity calculation sample inspection model of Ordovician bioturbated carbonate reservoirs in Tahe oilfield[J]. Acta Sedimentologica Sinica, 2021, 39(2): 482-492. [24] Meadows P S, Tait J. Modification of sediment permeability and shear strength by two burrowing invertebrates[J]. Marine Biology, 1989, 101(1): 75-82. [25] Perret J, Prasher S O, Kantzas A, et al. Three‐dimensional quantification of macropore networks in undisturbed soil cores[J]. Soil Science Society of America Journal, 1999, 63(6): 1530-1543. [26] Pierret A, Capowiez Y, Belzunces L, et al. 3D reconstruction and quantification of macropores using X-ray computed tomography and image analysis[J]. Geoderma, 2002, 106(3/4): 247-271. [27] Bastardie F, Capowiez Y, de Dreuzy J R, et al. X-ray tomographic and hydraulic characterization of burrowing by three earthworm species in repacked soil cores[J]. Applied Soil Ecology, 2003, 24(1): 3-16. [28] Tonkin N S, McIlroy D, Meyer R, et al. Bioturbation influence on reservoir quality: A case study from the Cretaceous Ben Nevis Formation, Jeanne d'Arc Basin, offshore Newfoundland, Canada[J]. AAPG Bulletin, 2010, 94(7): 1059-1078. [29] Qi Y A, Wang M, Zheng W, et al. Calcite cements in burrows and their influence on reservoir property of the Donghe sandstone, Tarim Basin, China[J]. Journal of Earth Science, 2012, 23(2): 129-141. [30] Adepehin E J, Bankole O M, Arifin M H. Poro-perm evolution in Oligo-Miocene coastal sandstones: Constraining the relative influence of sedimentary facies, mineralogy, and diagenesis on analogue reservoir quality of the Nyalau Formation, Borneo[J]. Marine and Petroleum Geology, 2022, 139: 105589. [31] Adam A, Swennen R, Abdulghani W, et al. Reservoir heterogeneity and quality of Khuff carbonates in outcrops of central Saudi Arabia[J]. Marine and Petroleum Geology, 2018, 89: 721-751. [32] 牛永斌,钟建华,钟福平,等. 柴达木盆地南缘石炭系Chondrites遗迹化石的发现及其地质意义[J]. 古地理学报,2008,10(5):529-535. Niu Yongbin, Zhong Jianhua, Zhong Fuping, et al. Discovery of trace fossil Chondrites in the Carboniferous in south margin of Qaidam Basin and its geological significance[J]. Journal of Palaeogeography, 2008, 10(5): 529-535. [33] 董小波,牛永斌. 成岩作用对豫西北马家沟组三段遗迹化石充填物孔隙发育的影响[J]. 海相油气地质,2015,20(3):17-27. Dong Xiaobo, Niu Yongbin. Diagenesis and effect of trace fossil fillings on pore development in Lower Ordovician Majiagou member-3 limestone in the northwest of Henan[J]. Marine Origin Petroleum Geology, 2015, 20(3): 17-27. [34] 张文凯,施泽进,田亚铭,等. 川东南志留系小河坝组致密砂岩孔隙类型及成因[J]. 岩性油气藏,2021,33(4):10-19. Zhang Wenkai, Shi Zejin, Tian Yaming, et al. Pore types and genesis of tight sandstone of Silurian Xiaoheba Formation in southeastern Sichuan Basin[J]. Lithologic Reservoirs, 2021, 33(4): 10-19. [35] Baniak G M, La Croix A D, Polo C A, et al. Associating X-ray microtomography with permeability contrasts in bioturbated media[J]. Ichnos, 2014, 21(4): 234-250. [36] Corlett H J, Jones B. Petrographic and geochemical contrasts between calcite-and dolomite-filled burrows in the Middle Devonian Lonely Bay Formation, Northwest Territories, Canada: Implications for dolomite formation in Paleozoic burrows[J]. Journal of Sedimentary Research, 2012, 82(9): 648-663. [37] 胡亚洲,牛永斌,崔胜利,等. 碳酸盐岩中生物潜穴充填特征及其诱导孔隙演化规律:以豫西奥陶系马家沟组三段为例[J]. 沉积学报,2019,37(4):690-701. Hu Yazhou, Niu Yongbin, Cui Shengli, et al. Characteristics of filled burrows in carbonates and the evolution of burrow-mediated pores: A case study from the Third member of the Ordovician Majiagou Formation, western Henan province[J]. Acta Sedimentologica Sinica, 2019, 37(4): 690-701. [38] 李峰峰,郭睿,刘立峰,等. 伊拉克M油田白垩系Mishrif组生物碎屑灰岩储集层非均质性成因[J]. 地球科学与环境学报,2020,42(3):297-312. Li Fengfeng, Guo Rui, Liu Lifeng, et al. Heterogeneity genesis of bioclastic limestone reservoirs of Cretaceous Mishrif Formation in M oilfield, Iraq[J]. Journal of Earth Sciences and Environment, 2020, 42(3): 297-312. [39] James N P, Choquette P W. Paleokarst[M]. New York: Springer Science & Business Media, 2012. [40] Loucks R G. Paleocave carbonate reservoirs: Origins, burial-depth modifications, spatial complexity, and reservoir implications[J]. AAPG Bulletin, 1999, 83(11): 1795-1834. [41] Li J, Zhang W Z, Luo X, et al. Paleokarst reservoirs and gas accumulation in the Jingbian field, Ordos Basin[J]. Marine and Petroleum Geology, 2008, 25(4/5): 401-415. [42] Rahimpour-Bonab H, Mehrabi H, Navidtalab A, et al. Palaeo-exposure surfaces in Cenomanian-Santonian carbonate reservoirs in the Dezful embayment, SW Iran[J]. Journal of Petroleum Geology, 2013, 36(4): 335-362. [43] Yang X F, Wang X Z, Tang H, et al. The Early Hercynian paleo-karstification in the Block 12 of Tahe oilfield, northern Tarim Basin, China[J]. Carbonates and Evaporites, 2014, 29(3): 251-261. [44] Tian F, Jin Q, Lu X B, et al. Multi-layered Ordovician paleokarst reservoir detection and spatial delineation: A case study in the Tahe oilfield, Tarim Basin, western China[J]. Marine and Petroleum Geology, 2016, 69: 53-73. [45] 孙瑞,陈曦,明爽,等. 鄂尔多斯盆地西北部奥陶系马家沟组斑状白云岩成因机理及储集特征[J]. 新疆地质,2012,30(4):442-446. Sun Rui, Chen Xi, Ming Shuang, et al. Formation mechanism and its reservoir characteristics of Ordovician Majiagou Formation mottled dolomite in northwestern Ordos Basin[J]. Xinjiang Geology, 2012, 30(4): 442-446. [46] Moore C H, Wade W J. Carbonate reservoirs: Porosity and diagenesis in a sequence stratigraphic framework[M]. 2nd ed. Amsterdam: Elsevier Science, 2013. [47] Rameil N. Early diagenetic dolomitization and dedolomitization of Late Jurassic and Earliest Cretaceous platform carbonates: A case study from the Jura Mountains (NW Switzerland, E France)[J]. Sedimentary Geology, 2008, 212(1/2/3/4): 70-85. [48] 沈瑛楚,宋新民,刘波,等. 伊拉克AD油田上白垩统Kh2段生物扰动与储层非均质性[J]. 天然气地球科学,2019,30(12):1755-1770. Shen Yingchu, Song Xinmin, Liu Bo, et al. Bioturbation and reservoir heterogeneity study of Upper Cretaceous Kh2 member, AD oilfield, Iraq[J]. Natural Gas Geoscience, 2019, 30(12): 1755-1770. [49] Alibrahim A, Duane M J, Dittrich M. Dolomite genesis in bioturbated marine zones of an Early-Middle Miocene coastal mud volcano outcrop (Kuwait)[J]. Scientific Reports, 2021, 11(1): 6636. [50] Rashid F, Glover P W J, Lorinczi P, et al. Porosity and permeability of tight carbonate reservoir rocks in the north of Iraq[J]. Journal of Petroleum Science and Engineering, 2015, 133: 147-161. [51] 许晴旸,范若颖,龚一鸣. 海相遗迹化石对显生宙生物大辐射事件的响应[J]. 古地理学报,2023,25(2):431-450. Xu Qingyang, Fan Ruoying, Gong Yiming. Marine ichnofossils as a record of major biodiversification events in the Phanerozoic[J]. Journal of Palaeogeography, 2023, 25(2): 431-450. [52] Ben-Awuah J, Eswaran P. Effect of bioturbation on reservoir rock quality of sandstones: A case from the Baram Delta, offshore Sarawak, Malaysia[J]. Petroleum Exploration and Development, 2015, 42(2): 223-231. [53] 李慧敏,赵振宇,宋微,等. 鄂尔多斯盆地西部奥陶系克里摩里组有效储层发育控制因素[J]. 石油实验地质,2023,45(3):434-442. Li Huimin, Zhao Zhenyu, Song Wei, et al. Controlling factors for effective reservoir development of Ordovician Kelimoli Formation in western Ordos Basin[J]. Petroleum Geology & Experiment, 2023, 45(3): 434-442. [54] Pemberton S G, Gingras M K. Classification and characterizations of biogenically enhanced permeability[J]. AAPG Bulletin, 2005, 89(11): 1493-1517. [55] Gingras M K, Pemberton S G, Henk F, et al. Applications of ichnology to fluid and gas production in hydrocarbon reservoirs[M]//MacEachern J A, Bann K L, Gingras M K, et al. Applied ichnology. SEPM Society for Sedimentary Geology, 2007. [56] Gingras M K, Baniak G, Gordon J, et al. Porosity and permeability in bioturbated sediments[J]. Developments in Sedimentology, 2012, 64: 837-868. [57] Gingras M K, Mendoza C A, Pemberton S G. Fossilized worm burrows influence the resource quality of porous media[J]. AAPG Bulletin, 2004, 88(7): 875-883. [58] Knaust D, Dorador J, Rodríguez-Tovar F J. Burrowed matrix powering dual porosity systems: A case study from the Maastrichtian chalk of the Gullfaks Field, Norwegian North Sea[J]. Marine and Petroleum Geology, 2020, 113: 104158. [59] Bednarz M, McIlroy D. Effect of phycosiphoniform burrows on shale hydrocarbon reservoir quality[J]. AAPG Bulletin, 2012, 96(10): 1957-1980. [60] Gilman J R, Kazemi H. Improved calculations for viscous and gravity displacement in matrix blocks in dual-porosity simulators (includes associated papers 17851, 17921, 18017, 18018, 18939, 19038, 19361 and 20174)[J]. Journal of Petroleum Technology, 1988, 40(1): 60-70. [61] Eltom H A, Alqubalee A, Yassin M A. Potential overlooked bioturbated reservoir zones in the shallow marine strata of the Hanifa Formation in central Saudi Arabia[J]. Marine and Petroleum Geology, 2021, 124: 104798. [62] Dorador J, Rodríguez-Tovar F J. High-resolution image treatment in ichnological core analysis: Initial steps, advances and prospects[J]. Earth-Science Reviews, 2018, 177: 226-237. [63] Dreyer T, Scheie Å, Walderhaug O. Minipermeameter-based study of permeability trends in channel sand bodies[J]. AAPG Bulletin, 1990, 74(4): 359-374. [64] Golab J A, Smith J J, Clark A K, et al. Effects of Thalassinoides ichnofabrics on the petrophysical properties of the Lower Cretaceous Lower Glen Rose Limestone, Middle Trinity Aquifer, northern Bexar County, Texas[J]. Sedimentary Geology, 2017, 351: 1-10. [65] Alqubalee A, Muharrag J, Salisu A M, et al. The negative impact of Ophiomorpha on reservoir quality of channelized deposits in mixed carbonate siliciclastic setting: The case study of the Dam Formation, Saudi Arabia[J]. Marine and Petroleum Geology, 2022, 140: 105666. [66] Eltom H A, Yassin M A, Cline J, et al. Incorporating outcrop observations and laboratory measurements in 3D reservoir models: An integrated study to enhance the workflow of modeling bioturbated carbonate reservoirs[J]. Marine and Petroleum Geology, 2023, 152: 106259. [67] Eltom H A, Rankey E C, Hasiotis S T, et al. Effect of bioturbation on petrophysical properties: Insights from geostatistical and flow simulation modeling[J]. Marine and Petroleum Geology, 2019, 104: 259-269. [68] La Croix A D, Gingras M K, Dashtgard S E, et al. Computer modeling bioturbation: The creation of porous and permeable fluid-flow pathways[J]. AAPG Bulletin, 2012, 96(3): 545-556. [69] Eltom H A, Goldstein R H. Use of geostatistical modeling to improve the understanding of permeability upscaling in isotropic and anisotropic burrowed reservoirs[J]. Marine and Petroleum Geology, 2021, 129: 105067. [70] Cunningham K J, Sukop M C, Curran H A. Carbonate aquifers[J]. Developments in Sedimentology, 2012, 64: 869-896. [71] Hsieh A I, Allen D M, MacEachern J A. Upscaling permeability for reservoir-scale modeling in bioturbated, heterogeneous tight siliciclastic reservoirs: Lower Cretaceous Viking Formation, Provost Field, Alberta, Canada[J]. Marine and Petroleum Geology, 2017, 88: 1032-1046. [72] Cunningham K J, Sukop M C, Huang H B, et al. Prominence of ichnologically influenced macroporosity in the karst Biscayne aquifer: Stratiform “super-K” zones[J]. GSA Bulletin, 2009, 121(1/2): 164-180. [73] Eltom H A, Alqubalee A M, Sultan A S, et al. Understanding the permeability of burrow-related gas reservoirs through integrated laboratory techniques[J]. Journal of Natural Gas Science and Engineering, 2021, 90: 103917. [74] Baniak G M, La Croix A D, Gingras M K. Recent advancements in characterizing permeability and porosity distributions in bioturbated flow media[J]. Earth-Science Reviews, 2022, 232: 104162. [75] La Croix A D, Gingras M K, Pemberton S G, et al. Biogenically enhanced reservoir properties in the Medicine Hat gas field, Alberta, Canada[J]. Marine and Petroleum Geology, 2013, 43: 464-477. [76] Webber K J, van Geuns L C. Framework for constructing clastic reservoir simulation models[J]. Journal of Petroleum Technology, 1990, 42(10): 1248-1297. [77] Muskat M. The flow of homogeneous fluids through porous media[J]. Soil Science, 1938, 46(2): 169. [78] Freeze R A, Cherry J A. Groundwater[M]. Englewood Cliffs: Prentice-Hall, 1979: 262-265. [79] Gingras M K, Pemberton S G, Mendoza C A, et al. Assessing the anisotropic permeability of Glossifungites surfaces[J]. Petroleum Geoscience, 1999, 5(4): 349-357. [80] Baniak G M, Gingras M K, Burns B A, et al. Petrophysical characterization of bioturbated sandstone reservoir facies in the Upper Jurassic Ula Formation, Norwegian North Sea, Europe[J]. Journal of Sedimentary Research, 2015, 85(1): 62-81. [81] MacEachern J A, Raychaudhuri I, Pemberton S G. Stratigraphic applications of the Glossifungites ichnofacies: Delineating discontinuities in the rock record[M]//Pemberton S G. Applications of ichnology to petroleum exploration: A core workshop. Tulsa: SEPM Core Workshop, 1992. [82] Pemberton S G, Frey R W. The Glossifungites ichnofacies: Modern examples from the Georgia coast, U.S.A.[M]//Curran H A. Biogenic structures: Their use in interpreting depositional environments. Tulsa: SEPM Special Publication, 1985. [83] Gingras M K, Pemberton S G, Saunders T. Bathymetry, sediment texture, and substrate cohesiveness; their impact on modern Glossifungites trace assemblages at Willapa Bay, Washington[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2001, 169(1/2): 1-21. [84] King M R, Botterill S E, Gingras M K, et al. Freshwater to low salinity expression of Cretaceous Glossifungites-demarcated autogenic stratigraphic surfaces, central Utah[J]. Ichnos, 2022, 29(1): 1-10. [85] Eltom H A, Syahputra M R N, El-Husseiny A, et al. Spatial complexity of burrow attributes and their impact on porosity and permeability distributions in bioturbated reservoirs[J]. Sedimentary Geology, 2023, 450: 106395. [86] Ingram M F. Basin analysis and hydrocarbon potential of the Bali Basin with expanded geothermal heat flow map of Indonesia[D]. Muncie: Ball State University, 2018. [87] Noble R A, Henk Jr F H. Hydrocarbon charge of a bacterial gas field by prolonged methanogenesis: An example from the East Java Sea, Indonesia[J]. Organic Geochemistry, 1998, 29(1/2/3): 301-314. [88] Lemiski R T, Hovikoski J, Pemberton S G, et al. Sedimentological ichnological and reservoir characteristics of the low-permeability, gas-charged Alderson member (Hatton gas field, southwest Saskatchewan): Implications for resource development[J]. Bulletin of Canadian Petroleum Geology, 2011, 59(1): 27-53. [89] Gingras M K, MacMillan B, Balcom B J, et al. Using magnetic resonance imaging and petrographic techniques to understand the textural attributes and porosity distribution in Macaronichnus-burrowed sandstone[J]. Journal of Sedimentary Research, 2002, 72(4): 552-558. [90] Dafoe L T, Gingras M K, Pemberton S G. Determining Euzonus mucronata burrowing rates with application to ancient Macaronichnus segregatis trace-makers[J]. Ichnos, 2008, 15(2): 78-90. [91] Ali S, Gingras M K, Wilson B, et al. The influence of bioturbation on reservoir quality: Insights from the Columbus Basin, offshore Trinidad[J]. Marine and Petroleum Geology, 2023, 147: 105983. [92] Gordon J B, Pemberton S G, Gingras M K, et al. Biogenically enhanced permeability: A petrographic analysis of Macaronichnus segregatus in the Lower Cretaceous Bluesky Formation, Alberta, Canada[J]. AAPG Bulletin, 2010, 94(11): 1779-1795. [93] Pemberton S G, MacEachern J A, Gingras M K, et al. Biogenic chaos: Cryptobioturbation and the work of sedimentologically friendly organisms[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2008, 270(3/4): 273-279. [94] Abdel-Fattah Z A, Gingras M K, Pemberton S G. Significance of hypoburrow nodule formation associated with large biogenic sedimentary structures in open-marine bay siliciclastics of the Upper Eocene Birket Qarun Formation, Wadi El-Hitan, Fayum, Egypt[J]. Sedimentary Geology, 2011, 233(1/2/3/4): 111-128. [95] Trabelsi A, Beg M A. Characterization and mapping of burrowed and microporous intervals in the Arab D Reservoir, Dukhan Field, Qatar[C]//Proceedings of the Abu Dhabi international petroleum exhibition and conference. Abu Dhabi: SPE, 2000. [96] Niu Y B, Cheng M Y, Zhang L J, et al. Bioturbation enhanced petrophysical properties in the Ordovician carbonate reservoir of the Tahe oilfield, Tarim Basin, NW China[J]. Journal of Palaeogeography, 2022, 11(1): 31-51. [97] Eltom H A, González L A, Alqubalee A, et al. Evidence for the development of a superpermeability flow zone by bioturbation in shallow marine strata, Upper Jubaila Formation, central Saudi Arabia[J]. Marine and Petroleum Geology, 2020, 120: 104512. [98] Kendall A C. Origin of dolomite mottling in Ordovician limestones from Saskatchewan and Manitoba[J]. Bulletin of Canadian Petroleum Geology, 1977, 25(3): 480-504. [99] Kendall A C. Anhydrite replacements of gypsum (satin-spar) veins in the Mississippian caprocks of southeastern Saskatchewan[J]. Canadian Journal of Earth Sciences, 1975, 12(7): 1190-1195. [100] Jin J S, Harper D A T, Rasmussen J A, et al. Late Ordovician massive-bedded Thalassinoides ichnofacies along the palaeoequator of Laurentia[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2012, 367-368: 73-88. [101] 刘航宇,田中元,刘波,等. 中东地区巨厚强非均质碳酸盐岩储层分类与预测:以伊拉克W油田中白垩统Mishrif组为例[J]. 石油学报,2019,40(6):677-691. Liu Hangyu, Tian Zhongyuan, Liu Bo, et al. Classification and prediction of giant thick strongly heterogeneous carbonate reservoirs in the Middle East area: A case study of Mid-Cretaceous Mishrif Formation in the W oilfield of Iraq[J]. Acta Petrolei Sinica, 2019, 40(6): 677-691. [102] 胡斌,齐永安,宋慧波,等. 中国遗迹学研究十年进展[J]. 古地理学报,2021,23(2):284-320. Hu Bin, Qi Yong’an, Song Huibo, et al. Research progress of Chinese ichnology in recent ten years[J]. Journal of Palaeogeography (Chinese Edition), 2021, 23(2): 284-320. [103] 牛永斌,齐永安,胡斌,等. 遗迹组构的精细分析功能及其应用:第15届国际遗迹组构专题研讨会综述[J]. 古地理学报,2019,21(5):767-782. Niu Yongbin, Qi Yong’an, Hu Bin, et al. Fine analysis functions and their application of ichnofabric: Outline of the 15th International Ichnofabric Workshop[J]. Journal of Palaeogeography (Chinese Edition), 2019, 21(5): 767-782. [104] Knaust D. Characterisation of a Campanian deep-sea fan system in the Norwegian Sea by means of ichnofabrics[J]. Marine and Petroleum Geology, 2009, 26(7): 1199-1211. [105] Reolid J, Betzler C. Ichnofabric logs for the characterization of the organic content in carbonates[J]. Marine and Petroleum Geology, 2018, 95: 246-254. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 353
- HTML全文浏览量: 71
- PDF下载量: 69
- 被引次数: 0

