

 下载:
下载:

-
生命塑造了宜居的地球,这其中的关键是被称为生命引擎的微生物[1]。众所周知,生命与矿物表现出明显的协同演化关系,导致矿物由最初的400多种发展到4 400多种,并表现出明显的阶段性[2]。岩石是由矿物组成的,它与微生物又会是怎样的一种关系?虽然岩石种类繁多,包括沉积岩、岩浆岩和变质岩,但至少沉积岩与微生物可能具有协同演化关系。
自从生命起源以后,微生物在地表环境无处不在,它势必影响在地表环境形成的沉积物和沉积岩。实际上,在地球表层系统发生的风化侵蚀作用、搬运作用、沉积成岩作用,无不贯穿着微生物的地质作用。在沉积物固结成岩的最关键阶段,水—沉积物界面的氧化—还原分带实际上就是根据微生物作用划分为氧化阶段、锰还原阶段、硝酸盐还原阶段、铁还原阶段、硫酸盐还原阶段、甲烷形成阶段。也就是说,水体环境的所有沉积物都经过了不同微生物功能群的洗礼之后才形成!所以从微生物分布情况、沉积物形成的环境等方面分析,微生物与沉积岩应该具有协同演化的关系。
-
地球系统过程涉及物理过程、化学过程和生物过程,其中表层系统的物理和化学过程都或多或少地有生物的参与,因此在表层系统形成的沉积物和沉积岩实际上是离不开生物作用的。关键在于生物参与的程度有多大、贡献有多大。沉积岩可分为自生沉积岩和他生沉积岩,许多自生沉积岩都有微生物作用的烙印。随着研究的深入,原来被认为他生沉积岩中的“碎屑岩”,其中的一些可能也是微生物参与形成的自生沉积岩。
-
许多灰岩无疑与生物或生物化学作用有关,特别是微生物的作用。海洋中的Ca2+要以碳酸盐形式沉淀下来,必须在碱性环境条件下,而微生物作用可以造成碱性环境条件。在海洋表层,由于光合自养微生物功能群的光合作用,大量消耗水体的CO2,导致海水表层的碱度增大,碳酸钙过饱和而沉淀下来。在浅水环境,许多底栖光合自养微生物直接形成了种类繁多的钙质微生物岩,包括叠层石、树形石、凝块石等[3],甚至一些大型藻类被直接钙化。当然,在远洋,海水表层沉淀的这些碳酸钙最后能否沉积,还与海洋的碳酸盐补偿深度(Carbonate Compensation Depth,CCD)相关。但无论如何,海洋的大量碳酸钙沉积与光合自养微生物功能群的作用有关。
近些年来,人们发现越来越多的碳酸钙不是从海水表面沉淀下来的,而是直接在海底的水—沉积物界面或者沉积物内部孔隙水里沉淀形成的,这些碳酸钙也被称为自生成因[4]或者海底碳酸盐工厂产物[5]。一般而言,在水—沉积物界面附近,随着一些有机质的氧化分解而出现一些有机酸,会使局部环境变成酸性,难以沉淀碳酸钙。但是,当有机酸彻底分解,随着埋深加大和氧化还原电位的进一步变化,许多化能自养微生物作用开始形成碱性的微环境。例如,铁还原过程、硫酸盐还原过程均可提高环境碱度。因此,碳酸钙就会在沉积物的孔隙水里因过饱和而沉淀,从而形成自生碳酸盐岩。
特别是,硫酸盐还原与甲烷氧化往往耦合在一起,从而在硫酸盐还原带和其下的甲烷形成带之间存在一个过渡带。在这个过渡带,经常出现碳酸钙的过饱和而形成许多结核状的碳酸钙。在华南埃迪卡拉纪陡山沱组的黑色岩系里,出现了大量巨大的碳酸钙结核,同时存在许多黄铁矿,很可能与这种微生物作用过程相关。
除了光合自养微生物和化能自养微生物分别形成的这两类碳酸钙以外,还有许多具有粒屑结构的碳酸钙,也可能与微生物作用有关。例如,一些鲕粒灰岩就与藻类或者细菌等微生物及其有机质的作用有关[6⁃8]。在二叠纪—三叠纪之交,许多鲕粒灰岩的鲕粒特别大,称为巨鲕,它们与微生物岩伴生[9]。
-
与灰岩相比,白云岩的成因更加复杂,因为它涉及钙离子和镁离子按照1∶1的比例进入晶格的难题,而且镁离子在海水中是以水合镁离子存在的,很难解离出来进入矿物晶格。虽然白云石的矿物成分和晶体结构极其简单,但其成因却非常复杂而成为白云石之谜。首先,现代海水对白云石是过饱和的,但却很少有沉淀。其次,地质时期白云岩分布十分广泛,但实验室在低温条件下难以无机合成白云石。因此,地质时期广泛分布的白云岩被认为是灰岩经过后期交代作用所致,或者是热化学成因。
自从1995年发现微生物可以沉淀白云石以来[10],白云岩的微生物成因引起了广泛关注。人们陆续发现了硫酸盐还原细菌、嗜盐古菌、产甲烷古菌、浮游自养微生物以及与微生物有关的一些有机分子或有机碎片都能沉淀白云石,从而建立了微生物白云岩的模式[11⁃12]。微生物之所以能够促进白云石的沉淀,主要是因为微生物表面具有一些活性官能团,这些官能团能从水合镁离子中解离出镁离子,从而在细胞表面形成白云石沉淀。
随着模拟实验发现越来越多的微生物能够沉淀出白云石,地质时期的白云岩也不断地被认为是微生物成因的。显生宙的二叠纪—三叠纪之交,海洋动物大灭绝之后,出现了白云岩事件,白云岩沉积从浅水地区向深水地区转移,被认为与微生物作用有关,特别是硫酸盐还原细菌的作用[13]。在元古宙埃迪卡拉纪,陡山沱组的盖帽白云岩被认为是低温环境的产物,而且很可能与微生物作用有关[14]。
需要指出的是,从实验模拟的微生物成因白云石到地质记录的微生物成因白云岩,还有很长的路要走,实验模拟与地质记录的衔接还没有完全建立起来。一是实验模拟的微生物成因白云石主要是无序白云石,而地质体中的白云岩都是有序白云石。二是没有建立起白云石微生物成因的识别标志。这两大难题在很大程度上困扰着人们对地质时期微生物成因白云岩的认识,亟待突破。
-
泥岩一直被认为是陆源碎屑岩,因而主要是物理成因,几乎不太可能将它与微生物建立起联系而具有生物成因。在水体环境,远离岸边依次出现砾岩、砂岩、粉砂岩和泥岩的分带就被认为是物理分选作用的产物。在海洋环境,泥岩外围更深水的地方往往沉积了生物成因的灰岩,那么离灰岩最近的泥岩是否也具有生物成因呢?
随着研究的深入,人们已经发现微生物成因泥岩的蛛丝马迹[15]。主要的依据有这样两方面。首先,微生物可以导致一些黏土矿物的相变或者直接沉淀黏土矿物。大家熟知的蒙脱石向伊利石转变,传统一直认为需要高温高压条件,因而需要在地下埋深数千米才能发生。然而,越来越多的实验模拟研究发现,沉积成岩作用带的几乎所有微生物功能群都能在常温常压下将蒙脱石转变为伊利石,因为微生物酶的催化效应与地质温度具有同样的效应[16]。微生物对黏土矿物的转变还形成了石英、长石、滑石等在高温环境常见的矿物[17]。许多泥岩里也出现了这些自形程度很高的不太可能是碎屑成因的矿物。所有这些黏土矿物以及其中的“碎屑”矿物均有可能是微生物成因。
特别值得注意的是,人们已经提出来,显生宙泥岩在志留纪以后的丰度明显增加,这可能与陆地植物的登陆有关。这些植物提供的有机质输入到水体对黏土矿物起絮凝作用,从而导致泥岩丰度在植物登陆后快速增加[18]。实际上,人们已经发现,在土壤系统,刚开始主要是高等植物输入的有机质,随着时间的增加,这些高等植物有机质逐渐被微生物利用而使土壤里微生物有机质越来越多,最后土壤有机质主要是由微生物来源的有机质构成[19]。结合这两方面的结果,微生物通过对陆地植物输入水体的有机质进行改造,这可能加强了对水体黏土矿物的絮凝作用,从而出现了更多的泥岩。同时,低等生物和高等植物登陆还增强了陆地成土作用,这在很大程度上也影响了黏土矿物工厂[20]。
尽管有许多迹象显示了微生物在地质时期泥岩形成中扮演了重要的角色,但到目前为止,还没有在地质记录中确认的微生物成因泥岩,这有待于后续的深入研究,特别是要建立微生物成因泥岩的识别标志,包括矿物组合、同位素组成特征以及有机分子特征等。许多泥岩往往含有大量的有机质,形成了黑色岩系,除了泥岩能吸附有机质以外,更可能是微生物成因的一个反映。
-
在自然界,铁的氧化还原作用往往与铁代谢微生物功能群有关,因而铁质岩具有微生物成因也就不足为奇了,甚至一些沉积型铁矿都与微生物的作用有关。在这方面比较典型的元古宙条带状硅铁建造(Banded Iron Formation,BIF),其中的磁铁矿就与铁代谢微生物在缺氧的海洋环境将Fe2+转变成Fe3+有关[21]。没有这些微生物的作用,很难想象Fe2+在缺氧环境下能被大量地氧化。除了BIF以外,地质时期出现的许多红层也可能与微生物作用有关,这些红层往往伴随缺氧的海洋环境里存在大量的还原性铁,并在随后的氧化环境被大量地氧化[22]。
磷是生命的必需元素,诸如磷块岩等磷质岩的微生物成因也就司空见惯了。磷元素在还原条件下容易活化而在氧化条件下沉淀,导致磷块岩经常出现在海洋的上升流区。在该区域,大量微生物作用使得底层水出现缺氧环境,磷得以活化,这些磷随后被上升流带入海水表层的氧化环境而沉淀,加之微生物对磷具有强大的吸附能力,大量的磷块岩沉积。磷块岩的沉积实际上就是一个微生物或者微生物化学过程。
如果说以上的铁质和磷质岩石的沉淀都与Eh驱动的微生物群落变化有关,那么硅质的沉淀则主要与pH有关。不同于钙质沉淀需要碱度较高的水体环境,硅质沉淀则需要在偏酸性的环境下沉淀。虽然无机过程可以导致硅质沉淀,特别是地质时期的热液喷口附近经常有硅质沉积,但地质时期的硅藻、放射虫等真核微生物则是硅质沉积的主要贡献者,特别是出现在海洋CCD以下的那些大量硅质堆积,赤道等低纬度地区主要与放射虫有关,而高纬度的海洋硅质则主要与硅藻有关。
-
正因为大部分的灰岩、白云岩、铁质岩、硅质岩和磷质岩主要是微生物成因的,一些泥岩也可能与微生物作用有关,因此地质时期的微生物与沉积岩应该具有协同演化的关系。特别是,由于微生物演化具有阶段性特征,气候环境的变化也具有明显的阶段性,从而导致微生物与沉积岩的协同演化也表现出明显的阶段性特征,使得不同地质时期出现不同类型的沉积岩。
-
太古宙生命刚开始起源,微生物的地质作用有限,以物理作用为主导。同时,这个时期地球表层系统总体是缺氧的环境条件。这些生物与环境特征影响了太古宙沉积岩的特征。
太古宙的矿物种类和岩石类型变化都不大,矿物主要为1 000~1 500种,岩石有几个重要特征(图1)。一是以火成岩为主,沉积岩相对比例较小,大部分时间沉积岩的体积占比在25%以下。二是沉积岩以杂砂岩为主。三是白云岩占有一定的比例,远比灰岩出现的时间要早,丰度也更高。四是条带状硅铁建造BIF在太古宙后期比较丰富,最近资料显示是整个地质时期丰度最高的时期[21]。后两个特征,特别是BIF主要与微生物的地质作用有关,尤其是缺氧环境条件下铁代谢微生物的作用。

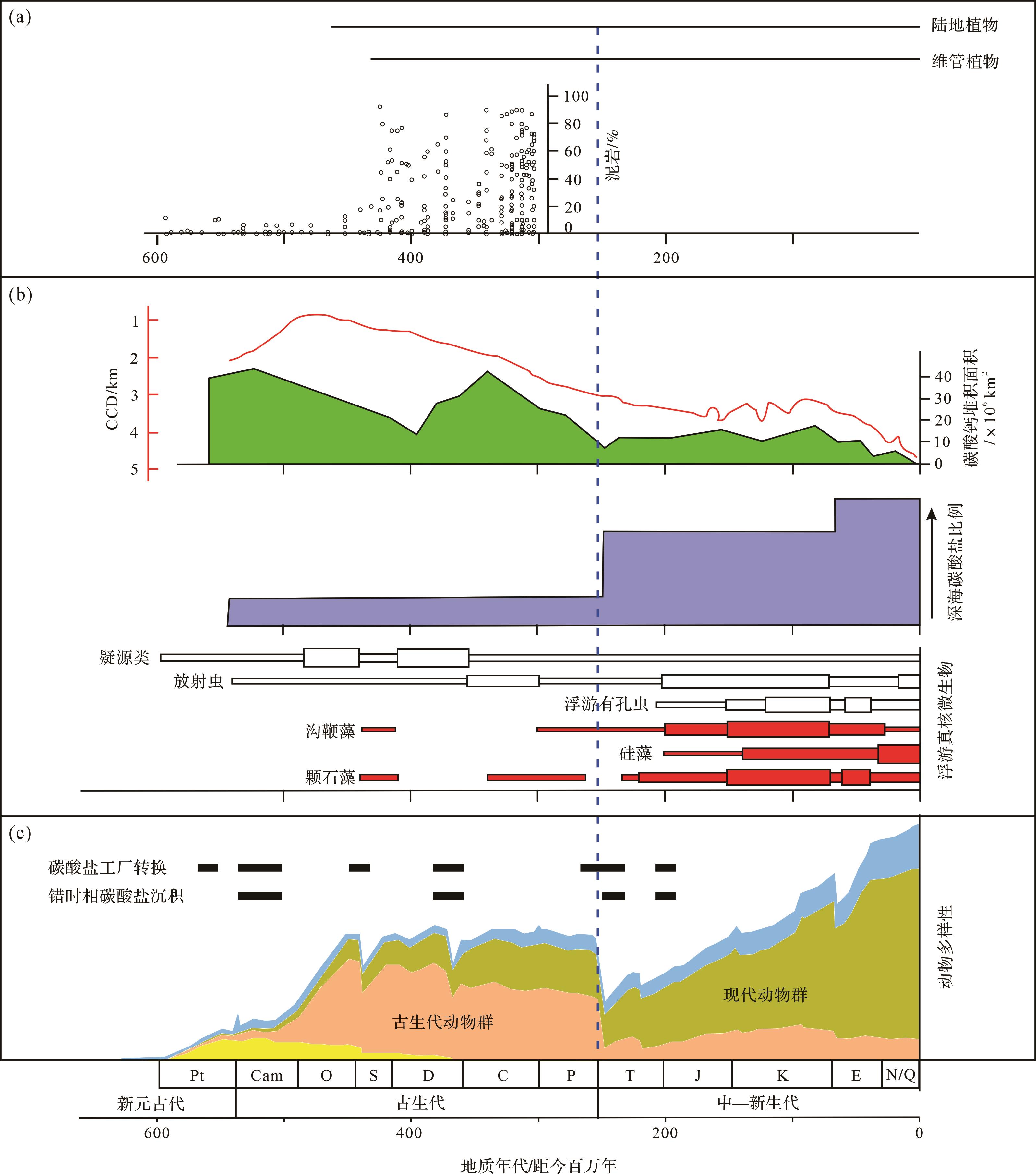
图 1 地球历史上生物、矿物、岩石和大气含氧量的变化
Figure 1. Evolution of organisms, minerals, rocks, and atmospheric oxygen level throughout Earth history
在太古宙早期,海相碳酸盐岩较少,呈不连续薄层分布于深水环境,主要形成于火山活动的宁静时期[26]。即使在太古宙中期,克拉通地区薄层浅水碳酸盐岩出现,厚度也不超过60 m[26]。但到了太古宙晚期,海相浅水碳酸盐岩分布广泛,厚度可达500 m,出现叠层石建造、颗粒岩等台地边缘沉积相带,台地边缘高差可达500 m[26],这些变化可能与微生物地质作用的增强有关。
-
随着产氧光合微生物乃至后期真核微生物的起源和爆发,元古宙的大气海洋环境出现巨大变化,从缺氧状态向氧化状态转变,矿物种类猛增,超过4 000种,沉积岩也发生了革命性的变化(图1),元古宙因而是一个微生物与环境之间发生强烈相互作用的关键时期。
首先,岩石的“水火格局”出现革命性变化。火成岩占比减少,沉积岩的相对丰度显著增加,到了后期可以占体积比的75%,基本接近显生宙的水平。这种“水火格局”与太古宙形成了鲜明的对比,可以说沉积岩石圈发生了革命性的变化。
其次,微生物成因的岩石大幅度增加。杂砂岩的相对比例降低,间接地反映了微生物成因的岩石在增加。最为典型的微生物成因岩石是叠层石,它的丰度是整个地质时期最高的,这与元古宙是蓝细菌的时代有关。这些蓝细菌形成了丰富多彩的微生物岩,除了各种各样的叠层石以外,还有树形石、凝块石等。可能与微生物作用有关的白云岩也越来越多,元古宙晚期成了地质时期丰度最高的时期。灰岩丰度也不断地增高。BIF在早元古代还存在,而且丰度较高,但到了中、新元古代就消失了。需要指出的是,元古宙微生物成因岩石的增加也伴随着微生物沉积构造(Microbially-Induced Sedimentary Structure,MISS)的多样性[27]。同时,微生物成因的碳酸盐台地在形态上也发生多样性变化,既有缓坡也有陡坡的形态[26,28]。这些特征均体现了微观尺度的微生物能够产生宏观尺度的效应。
特别是,前人对元古宙微生物成因碳酸盐岩有许多总结[26,28],当时稳定的克拉通因发育碳酸盐沉积的面积显著增加而形成了大套厚层碳酸盐岩。古元古代碳酸盐岩台地分布广泛,沉积相的水深差异已经与显生宙接近。先形成的可能是缓坡,接着形成镶边台地,台地边缘发育宽阔的叠层石礁而形成1 000 m的地势差异[26]。中元古代出现孤立叠层石建造,可能与碳酸盐生产率的不同进而形成不同的地形有关。新元古代,由于藻类出现,或者后生动物的破坏,叠层石丰度下降,导致浅水碳酸盐工厂的转变,从底栖微生物工厂演化为浮游微生物灰泥工厂[26]。
再次,一些“碎屑”岩石也开始增多。传统观点认为的碎屑岩石——长石砂岩、石英砂岩和泥岩(页岩)不断增多,这与超大陆的聚合、大陆地壳不断增生有关,是物理作用的体现。石英砂岩的相对比例增多,说明化学风化作用增强,可能也与微生物参与的风化作用有关。泥岩从古元古代到新元古代不断增加的趋势特别明显,尤其是在新元古代丰度非常高。Sr同位素、K/Al和黏土矿物分布等证据显示,在新元古代末期,大陆边缘泥岩丰度突然升高与黏土矿物工厂的变化有关[20]。当时陆地的地衣等低等生物开始繁盛,成土作用加强导致黏土矿物激增,并向海输送到大陆边缘而沉积下来。这些沉积于大陆边缘的泥岩同时埋藏了大量的有机碳,在一定程度上触发了第二次大氧化事件以及随后的动物起源和爆发。
最后,沉积岩出现硅质—磷质—钙质转变。在新元古代末期,呈现出硅质—磷质—钙质岩石或者生物壳体成分的转变,体现了海洋水化学成分的变化及其与生物相互作用的演变。中元古代乃至新元古代存在大量的硅化现象,甚至许多白云岩都被强烈硅化。之后,在新元古代晚期,出现许多磷块岩,甚至磷酸盐化的生物壳体。最后,出现了大量的钙质沉积和钙质壳体化石,包括著名的小壳化石。从硅质到磷质再到钙质的转变,可能与大气海洋环境的pH、Eh变化以及海水化学成分的变化有关,并叠加了生物(从原核微生物到真核微生物再到多细胞动物)的作用。早期的硅质沉积可能与海水偏酸性并以原核微生物为主导有关。磷质的出现反映了海洋环境条件出现频繁的氧化还原波动,并开始有真核微生物的参与。最后出现钙质,反映了海水碱度增大,并出现真核生物的参与,特别是多细胞动物壳体的钙化。
-
到了显生宙,地球表层系统彻底氧化,包括深海也出现氧化。多细胞动物出现爆发,并在生态系统占据主导地位,早古生代植物开始登陆。这些变化影响了沉积岩的演化。
首先,岩石的“水火格局”没有大变。显生宙沉积岩与火成岩的占比基本是从元古宙末期继承来的,没有发生很大的变化,大约是3:1的比例。显生宙最大的变化是各类沉积岩相对比例的变化(图1)。
其次,泥岩丰度剧增并呈现红黑之间的频繁交替。沉积岩最大的变化是泥岩的增多,可占地表所有岩石体积比的50%,尤其是在志留纪以后大幅度增加(图1、图2a)。这个丰度的剧增被认为与植物登陆有关[18]。如前所述,这可能是植物—微生物的相互作用通过有机质絮凝作用影响了黏土矿物的沉淀,植物登陆还改变了陆地成土作用进而加强了黏土矿物的形成。如果说泥岩丰度在新元古代末期的增加与陆地低等生物的发展而导致成土作用增强有关[20],那么志留纪泥岩丰度的激增则与高等植物大发展导致成土作用的增强有关,由此形成了两次黏土矿物工厂生产体制的升级。在这个大背景下,显生宙海相泥岩沉积序列还出现黑色岩系与红层的交替现象,这体现了海洋氧化还原条件的变化以及微生物功能群的变化[22]。

图 2 显生宙泥岩、碳酸盐岩与生物演化的协同关系
Figure 2. Coevolution of mudstones and carbonate rocks with organisms in the Phanerozoic
再次,碳酸盐工厂因生物与环境的变化而不断地转换(图2b)。第一,从总体上来看,白云岩占比在新元古代—寒武纪之交达到峰值后开始不断降低,但灰岩丰度在增加(图1),这反映了多细胞动物的作用在加强,而微生物的作用开始减弱。第二,从古生代到中—新生代,碳酸盐沉积从浅水地区不断向深水地区转移。古生代出现大量的多细胞动物产生的骨架灰岩或者大量宏观底栖藻类的钙化,这些主要出现在浅水地区。在古—中生代生物大灭绝后,出现了现代海洋占优势的三大红枝藻系,分别为硅藻、颗石藻和沟鞭藻[29],颗石藻等使得碳酸钙不断地向深水环境输送(图2b),由此出现著名的中中生代革命,海洋水化学发生变化[30]。
最后,在显生宙一些重大地质突变期出现了许多错时相(anachronistic facies)碳酸盐沉积,碳酸盐工厂发生多次变化(图2c)。这些沉积一般仅限于前寒武纪的正常浅海环境或者在极端环境才出现,但在显生宙一些生物大灭绝后的残存期在正常浅海环境短暂地“复辟”了,成为“时代上错位的相”。这些错时相包括微生物岩、鲕粒灰岩、蠕虫状灰岩、同生扁平砾石灰岩、薄层泥晶灰岩或者条带状灰岩、海底碳酸盐胶结扇等[5,34]。它们的沉积环境往往极度异常,形成于低(贫)氧浅海环境,缺乏正常宏体生物,保存原始薄—纹层状的层理,貌似深海沉积而实为浅水沉积(相对较深水),因此它们往往与各类微生物的作用有关。在显生宙,出现了多次从骨架灰岩向钙质微生物岩的碳酸盐工厂转变[32]。同时,一些MISS构造也广泛发育。
-
随着地质微生物研究的不断深入,人们发现越来越多的沉积岩具有微生物成因。条带状硅铁建造、灰岩、白云岩、泥岩以及其他硅质和磷质岩石都与微生物作用密切相关。许多沉积岩具有微生物成因,导致微生物与沉积岩具有协同演化的关系。太古宙以物理作用为主,而且以火成岩为主,但微生物作用形成的条带状硅铁建造是一大重要特征。元古宙的沉积岩比火成岩多,生物作用明显加强,大量钙质微生物岩出现,特别是叠层石是地质历史时期丰度最高的,白云岩也出现峰值。元古宙与显生宙转换时期出现了硅质、磷质到钙质的转换。在显生宙,白云岩丰度降低,碳酸盐工厂从骨架灰岩到钙质微生物岩的多次转换,植物登陆后泥炭在志留纪以后大量发育。沉积岩的丰度和种类在不同地质时期的变化与微生物的地史演化具有明显的关系。
微生物与沉积岩这些协同演化最终会对岩石圈乃至整个地球系统产生重要影响[35]。太古宙晚期出现的大量微生物成因的条带状含铁建造,具有高密度和大量铁组分,容易进入地幔而导致地幔的不均一性,并会引发火山活动[36]。这些微生物成因的BIF甚至被认为可能与板块运动的启动有关联[37]。在元古宙初期和末期,雪球地球的出现使得海洋里形成了大量沉积物,这些沉积物进入俯冲带而增强了润滑作用[38],导致前寒武纪两次构造活跃期和两次超大陆的聚合,分别形成了哥伦比亚和冈瓦纳超大陆[39]。在显生宙,红枝藻系的大发展,使得碳酸盐沉积从浅水向深水转移。深水碳酸盐沉积随着板块俯冲输入地幔深处,对深部碳循环产生影响[40⁃41]。一方面,在深部的金刚石里检测到来自地球表层系统与细菌有关的碳。另一方面,在一些板块俯冲带位置,深部地幔的碳特别富集。这都体现了微生物通过沉积物对深部过程产生的重要影响。
尽管人们对微生物与沉积岩的协同演化有了一些框架性认识,但目前还缺乏定量化的研究。一方面,各类沉积岩在地质时期的分布和丰度还需要大数据方面的定量化研究。另一方面,微生物成因的沉积岩更需要定量化的分析。前寒武纪的白云岩到底有多少与微生物作用有关,显生宙的泥岩又有多少是植物与微生物相互作用的产物,这些都有待于深入的定量化分析。微生物通过生物地球化学循环对深部过程和地球系统产生了重要影响,但微生物成因的沉积岩,通过哪些途径对地球系统产生影响,影响的深度和程度有多大,这些途径和影响又是如何随时间发生演变的,都有待于今后破解。
Coevolution of Microorganisms and Sedimentary Rocks
-
摘要: 意义 生物圈是地球区别于其他星球的一大关键,微生物因时空分布广、代谢功能多样、繁殖快、丰度高而对地球宜居性产生重要影响,探索微生物与沉积岩的协同演化关系对了解宜居地球的形成过程与途径具有重要意义。【 进展 】太古宙虽然以火成岩为主,少量的沉积岩也以物理作用为主,但铁代谢微生物广泛参与了条带状硅铁建造的形成。元古宙的沉积岩发生了革命性变化,其丰度远比火成岩多,而且微生物作用明显加强。元古宙出现了大量丰富多彩的钙质微生物岩和微生物成因沉积构造,白云岩出现峰值,微生物形成的碳酸盐台地形态也多变。在元古宙末期,泥岩丰度的增加与陆地低等生物繁盛而导致黏土矿物工厂的生产机制升级有关。在元古宙与显生宙的过渡时期,出现了从硅质、磷质到钙质的转换,这与原核微生物向真核微生物再向多细胞动物的演化以及海洋环境的pH和Eh等条件变化有关。到了显生宙,多细胞动物大发展,而微生物群落受到压制,白云岩丰度开始降低。在真核微生物方面,古生代绿藻类的优势被中—新生代的红枝藻系所代替,使得碳酸钙沉积从浅水向深水扩展。特别是,在显生宙一些重大突变期,碳酸盐工厂出现从骨架灰岩到钙质微生物岩的多次大转换,并且出现一些错时相和微生物成因沉积构造,这些都与宏体生物出现危机后微生物生态系统的短暂发展有关。泥岩丰度在志留纪以后猛增,这与高等植物登陆导致黏土矿物工厂的又一次变革以及植物—微生物相互作用导致有机质絮凝作用的加强有关。【 结论与展望 】地质时期的条带状硅铁建造、灰岩、白云岩、磷块岩、硅质岩等沉积岩都与微生物作用密切相关,甚至一部分泥岩的形成也可能与微生物作用有关。微生物参与了许多沉积岩的形成,导致微生物在地质时期与沉积岩具有协同演化的关系。未来可以利用地质大数据定量化研究各类沉积岩在地质时期的分布和丰度及其与不同微生物之间的关系,由此进一步探索地球深部与表层的联动。Abstract: Significance Earth differs from other planets because of the presence of the biosphere. Microorganisms in the biosphere have played, and are playing, important roles in shaping Earth habitability owing to their wide spatiotemporal occurrence, diverse metabolism functions, rapid multiplication, and great abundance. To explore the coevolution of microorganisms and sedimentary rocks is of great significance in understanding the processes and dynamics of the formation of Earth habitability. [ Progress ] The Archean is dominated by magmatic rocks, and its sedimentary rocks are primarily related to physical processes. However, banded iron formation is related to the contribution of photoferrotrophic bacteria. Significant changes occurred in the Proterozoic when sedimentary rocks overtook magmatic rocks in abundance and diverse biogenic rocks (calcimicrobialites and dolomites) greatly increased in abundance. Microbially-induced sedimentary structures were also diverse and occurred in great abundance. Mudstone began to increase in abundance at the end of the Proterozoic owing to the development of microorganisms on land, which generated a new clay factory. The transition interval across the Neoproterozoic and Cambrian boundary witnessed the shift from siliceous to phosphatic and finally to calcareous rocks, coincident with the evolution from prokaryotes to eukaryotic microorganisms and finally to metazoans, showing their causal relationship. In the Phanerozoic, dolomite exhibited a sharp decrease in abundance due to the metazoan expansion and its suppression on microorganisms. Green algae dominance in the Paleozoic shifted to the dominance of red algae in the Mesozoic and Cenozoic, favoring the expansion of calcium carbonate from shallow to deep water. In particular, carbonate factory showed the shift from skeleton limestones to calcimicrobialites for at least five times, in association with the occurrence of anachronistic facies as well as microbially-induced sedimentary structures; these are due to the transient expansion of microbial communities immediately after the biotic crisis. Mudstones increased in abundance immediately after the development of terrestrial ecosystems due to the innovation of a clay factory on land and the enhanced precipitation of clay minerals caused by organic-induced flocculation. [ Conclusions and Prospects ] Microbial involvement of sedimentation resulted in the biogenic formation of sedimentary rocks including banded iron formations, limestones, dolomites, phosphorites, siliceous rocks, and certain mudstones, which in turn have caused the coevolution of microbes and sedimentary rocks throughout Earth history. Current research has summarized such a co-evolution framework on the geological timescales. Future studies will quantitatively define the abundance of various sedimentary rocks over geological time based on reanalysis of the geological database, and quantitatively analyze microbial sedimentary rocks to better understand the coupled deep Earth-surface processes.
-
Key words:
- biopetrology /
- geomicrobiology /
- geobiology /
- lithosphere /
- biosphere /
- biogeological process
-
-
[1] 谢树成,焦念志,罗根明,等. 海洋生物碳泵的地质演化:微生物的碳汇作用[J]. 科学通报,2022,67(15):1715-1726. Xie Shucheng, Jiao Nianzhi, Luo Genming, et al. Evolution of biotic carbon pumps in Earth history: Microbial roles as a carbon sink in oceans[J]. Chinese Science Bulletin, 2022, 67(15): 1715-1726. [2] Hazen R M, Ferry J M. Mineral evolution: Mineralogy in the fourth dimension[J]. Elements, 2010, 6(1): 9-12. [3] 史晓颖,张传恒,蒋干清,等. 华北地台中元古代碳酸盐岩中的微生物成因构造及其生烃潜力[J]. 现代地质,2008,22(5):669-682. Shi Xiaoying, Zhang Chuanheng, Jiang Ganqing, et al. Microbial mats from the Mesoproterozoic carbonates of the North China Platform and their potential for hydrocarbon-generation[J]. Geoscience, 2008, 22(5): 669-682. [4] Schrag D P, Higgins J A, MacDonald F A, et al. Authigenic carbonate and the history of the global carbon cycle[J]. Science, 2013, 339(6119): 540-543. [5] Greene S E, Bottjer D J, Corsetti F A, et al. A subseafloor carbonate factory across the Triassic-Jurassic transition[J]. Geology, 2012, 40(11): 1043-1046. [6] 李飞,易楚恒,李红,等. 微生物成因鲕粒研究进展[J]. 沉积学报,2022,40(2):319-334. Li Fei, Yi Chuheng, Li Hong, et al. Recent advances in ooid microbial origin: A review[J] Acta Sedimentologica Sinica, 2022, 40(2): 319-334. [7] Riaz M, Banerjee S, Latif K, et al. Understanding the origin of ancient carbonate ooids: Recent findings[J]. International Geology Review, 2023, doi: 101080/00206814.2023.2220390 . [8] 郭芪恒,金振奎,史书婷,等. 鲕粒成因研究进展[J]. 沉积学报,2023,41(4):959-967. Guo Qiheng, Jin Zhenkui, Shi Shuting, et al. Research progress on the formation of ooids[J]. Acta Sedimentologica Sinica, 2023, 41(4): 959-967. [9] Li F, Yan J X, Chen Z Q, et al. Global oolite deposits across the Permian-Triassic boundary: A synthesis and implications for palaeoceanography immediately after the end-Permian biocrisis[J]. Earth-Science Reviews, 2015, 149: 163-180. [10] Vasconcelos C, McKenzie J A, Bernasconi S, et al. Microbial mediation as a possible mechanism for natural dolomite formation at low temperatures[J]. Nature, 1995, 377(6546): 220-222. [11] Qiu X, Wang H M, Yao Y C, et al. High salinity facilitates dolomite precipitation mediated by Haloferax volcanii DS52[J]. Earth and Planetary Science Letters, 2017, 472: 197-205. [12] Liu D, Yu N, Papineau D, et al. The catalytic role of planktonic aerobic heterotrophic bacteria in protodolomite formation: Results from lake Jibuhulangtu Nuur, Inner Mongolia, China[J]. Geochimica et Cosmochimica Acta, 2019, 263: 31-49. [13] Li M T, Song H J, Algeo T J, et al. A dolomitization event at the oceanic chemocline during the Permian-Triassic transition[J]. Geology, 2018, 46(12): 1043-1046. [14] Chang B, Li C, Liu D, et al. Massive formation of early diagenetic dolomite in the Ediacaran ocean: Constraints on the “dolomite problem”[J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(25): 14005-14014. [15] Konhauser K O, Urrutia M M. Bacterial clay authigenesis: A common biogeochemical process[J]. Chemical Geology, 1999, 161(4): 399-413. [16] 谢树成,刘邓,邱轩,等. 微生物与地质温压的一些等效地质作用[J]. 中国科学(D辑):地球科学,2016,46(8):1087-1094. Xie Shucheng, Liu Deng, Qiu Xuan, et al. Microbial roles equivalent to geological agents of high temperature and pressure in deep Earth[J]. Science China (Seri. D): Earth Sciences, 2016, 46(8): 1087-1094. [17] Liu D, Dong H L, Wang H M, et al. Low-temperature feldspar and illite formation through bioreduction of Fe(III)-bearing smectite by an alkaliphilic bacterium[J]. Chemical Geology, 2015, 406: 25-33. [18] McMahon W J, Davies N S. Evolution of alluvial mudrock forced by early land plants[J]. Science, 2018, 359(6379): 1022-1024. [19] Liang C, Zhu X F. The soil microbial carbon pump as a new concept for terrestrial carbon sequestration[J]. Science China Earth Sciences, 2021, 64(4): 545-558. [20] Kennedy M, Droser M, Mayer L M, et al. Late Precambrian oxygenation; Inception of the clay mineral factory[J]. Science, 2006, 311(5766): 1446-1449. [21] Konhauser K O, Planavsky N J, Hardisty D S, et al. Iron formations: A global record of Neoarchaean to Palaeoproterozoic environmental history[J]. Earth-Science Reviews, 2017, 172: 140-177. [22] Song H J, Jiang G Q, Poulton S W, et al. The onset of widespread marine red beds and the evolution of ferruginous oceans[J]. Nature Communications, 2017, 8(1): 399. [23] Veizer J, Mackenzie F T. Evolution of sedimentary rocks[J]. Treatise on Geochemistry, 2003, 7: 369-407. [24] Peters S E, Husson J M, Wilcots J. The rise and fall of stromatolites in shallow marine environments[J]. Geology, 2017, 45(6): 487-490. [25] Lyons T W, Reinhard C T, Planavsky N J. The rise of oxygen in Earth's early ocean and atmosphere[J]. Nature, 2014, 506(7488): 307-315. [26] Pomar L. Chapter 12, Carbonate systems[M]// Scarselli N, Adam J, Chiarella D, et al. Regional Geology and Tectonics (Second Edition). Elsevier, 2020: 235-311. [27] Noffke N, Knoll A H, Grotzinger J P. Sedimentary controls on the formation and preservation of microbial mats in siliciclastic deposits: A case study from the Upper Neoproterozoic Nama Group, Namibia[J]. Palaios, 2002, 17(6): 533-544. [28] Pomar L, Hallock P. Carbonate factories: A conundrum in sedimentary geology[J]. Earth-Science Reviews, 2008, 87(3/4): 134-169. [29] Falkowski P G, Katz M E, Knoll A H, et al. The evolution of modern eukaryotic phytoplankton[J]. Science, 2004, 305(5682): 354-360. [30] Ridgwell A. A Mid Mesozoic revolution in the regulation of ocean chemistry[J]. Marine Geology, 2005, 217(3/4): 339-357. [31] Eichenseer K, Balthasar U, Smart C W, et al. Jurassic shift from abiotic to biotic control on marine ecological success[J]. Nature Geoscience, 2019, 12(8): 638-642. [32] Chen Z Q, Tu C Y, Pei Y, et al. Biosedimentological features of major microbe-metazoan transitions (MMTs) from Precambrian to Cenozoic[J]. Earth-Science Reviews, 2019, 189: 21-50. [33] Sepkoski Jr J J. A kinetic model of Phanerozoic taxonomic diversity. III. Post-Paleozoic families and mass extinctions[J]. Paleobiology, 1984, 10(2): 246-267. [34] 赵小明,牛志军,童金南,等. 早三叠世生物复苏期的特殊沉积:“错时相”沉积[J]. 沉积学报,2010,28(2):314-323. Zhao Xiaoming, Niu Zhijun, Tong Jinnan, et al. The distinctive sediments in the Early Triassic recovery time: “anachronistic facies”[J]. Acta Sedimentologica Sinica, 2010, 28(2): 314-323. [35] 谢树成,罗根明,朱宗敏. 地球表层系统对深部圈层时空演变的影响[J/OL]. 科学通报,2023,doi: 10.1360/TB-2023-0002 . Xie Shucheng, Luo Genming, Zhu Zongmin. Surface system impact on the spatiotemporal evolution of deep Earth[J/OL]. Chinese Science Bulletin, 2023, doi: 10.1360/TB-2023-0002 . [36] Keller D S, Tassara S, Robbins L J, et al. Links between large igneous province volcanism and subducted iron formations[J]. Nature Geoscience, 2023, 16(6): 527-533. [37] Zhang S X, Li Y L, Leng W, et al. Photoferrotrophic bacteria initiated plate tectonics in the Neoarchean[J]. Geophysical Research Letters, 2023, 50(13): e2023GL103553. [38] Zeichner S S, Nghiem J, Lamb M P, et al. Early plant organics increased global terrestrial mud deposition through enhanced flocculation[J]. Science, 2021, 371(6528): 526-529. [39] Sobolev S V, Brown M. Surface erosion events controlled the evolution of plate tectonics on Earth[J]. Nature, 2019, 570(7759): 52-57. [40] Nisbet E G, Mattey D P, Lowry D. Can diamonds be dead bacteria?[J]. Nature, 1994, 367(6465): 694-697. [41] Müller R D, Mather B, Dutkiewicz A, et al. Evolution of Earth's tectonic carbon conveyor belt[J]. Nature, 2022, 605(7911): 629-639. -

 点击查看大图
点击查看大图

图(2)
计量
- 文章访问数: 608
- HTML全文浏览量: 127
- PDF下载量: 231
- 被引次数: 0

