
HTML
-
了解长时间尺度的植被演化过程对于研究现代植被的分布[1⁃2]、丰度[3⁃4]以及遗传基因结构[5⁃7]至关重要,古植被信息也为过去和未来的气候变化提供了重要的参考[8]。在古环境重建的研究中,需明确建立一些古气候代用指标来表征并重建当时的气候环境。传统的古植被重建代用指标主要有孢粉、植物大化石及植硅体等[9⁃11]。利用现代得到的孢粉数据建立环境因子转换函数,可以反演到沉积样品中实现古气候因子的定量重建[12⁃14],但此重建的基础主要是基于大区域的孢粉记录,在代表性和区域尺度上均有一定的局限性[15]。孢粉、植物大化石及植硅体并非是高分辨率且精确完整适用于任何类型沉积物的古植被代用指标,也并非所有植物信息都会以这3种形式进行保存,其中 “沉默”的一部分却无从得到[10,15]。
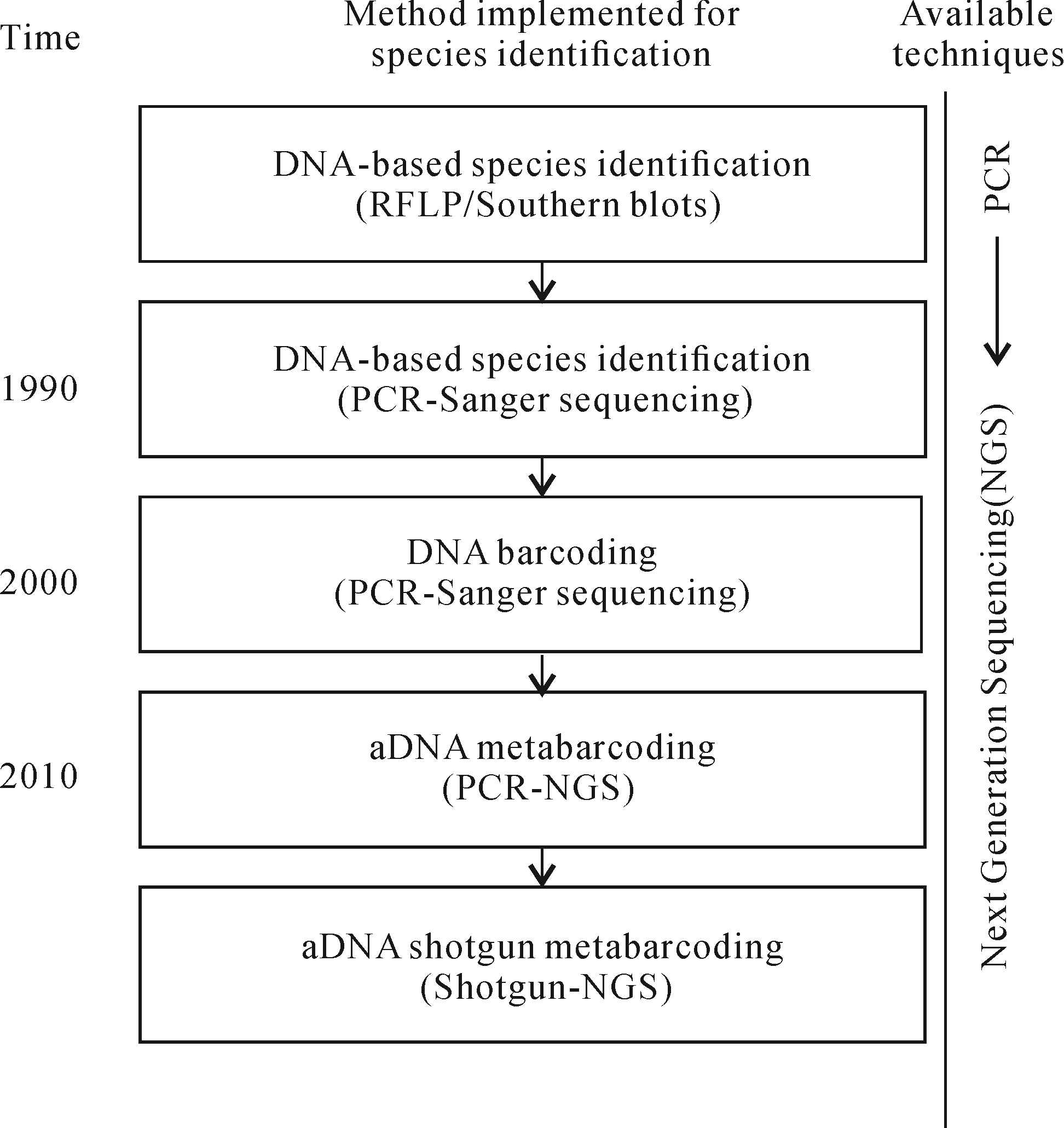
Willerslev et al. [16]在Science上的一篇关于提取到的西伯利亚和新西兰地区猛犸象及恐龙鸟的古DNA(Ancient DNA, aDNA)的报道,首次开启了分子生物学在古生态环境重建中应用的序幕。此后,分子学技术被广泛应用于众多古环境载体中,如冰川[17]、湖相沉积[18⁃20]及海洋沉积[21⁃23]等。研究表明,aDNA数据对其他古环境代用指标如孢粉、化石等是非常好的补充,是一种最新发展的有效物种鉴定技术。其目前最主流的分析方法是Metabarcoding(DNA条形码技术),已被广泛地应用于古生态学的研究,即从环境样品中提取DNA并结合第二代测序技术来实现对环境样品中所有存在物种的高精度检测,极大地推动了古生态研究的发展,已经成为目前国际古气候古植被研究的热点。因此,本文从沉积植物aDNA的概念、来源、保存、沉积过程、文献计量学统计、技术的发展应用、Metabarcoding及具体实验内容等方面进行了初步总结,并提出对未来研究方向的展望,为相关研究提供新的思路。
-
基于分子标记的种群遗传学研究揭示了第四纪气候演变对于现存植物基因结构的影响[24]。大量孢粉、植物大化石及基因方面的数据同时展示了末次冰期以来的植被分布[25⁃26]。古遗传学(如aDNA的研究)为我们研究古植被提供了一个非常有力的手段。古DNA研究主要为机体死亡经历水解和氧化后而保存的基因物质(DNA分子)[26⁃27]。在植物中,这些DNA分子主要赋存于植物残体组织,如枝干、果实和种子的厚壁组织及植物化石残体中[28],这些埋藏于沉积物中的古植物来源DNA就是沉积植物aDNA。沉积植物aDNA属于古环境DNA的范畴,是我们分析古植被信息的主要手段,也是近十年来应用非常广泛的古植被重建前沿方法。
-
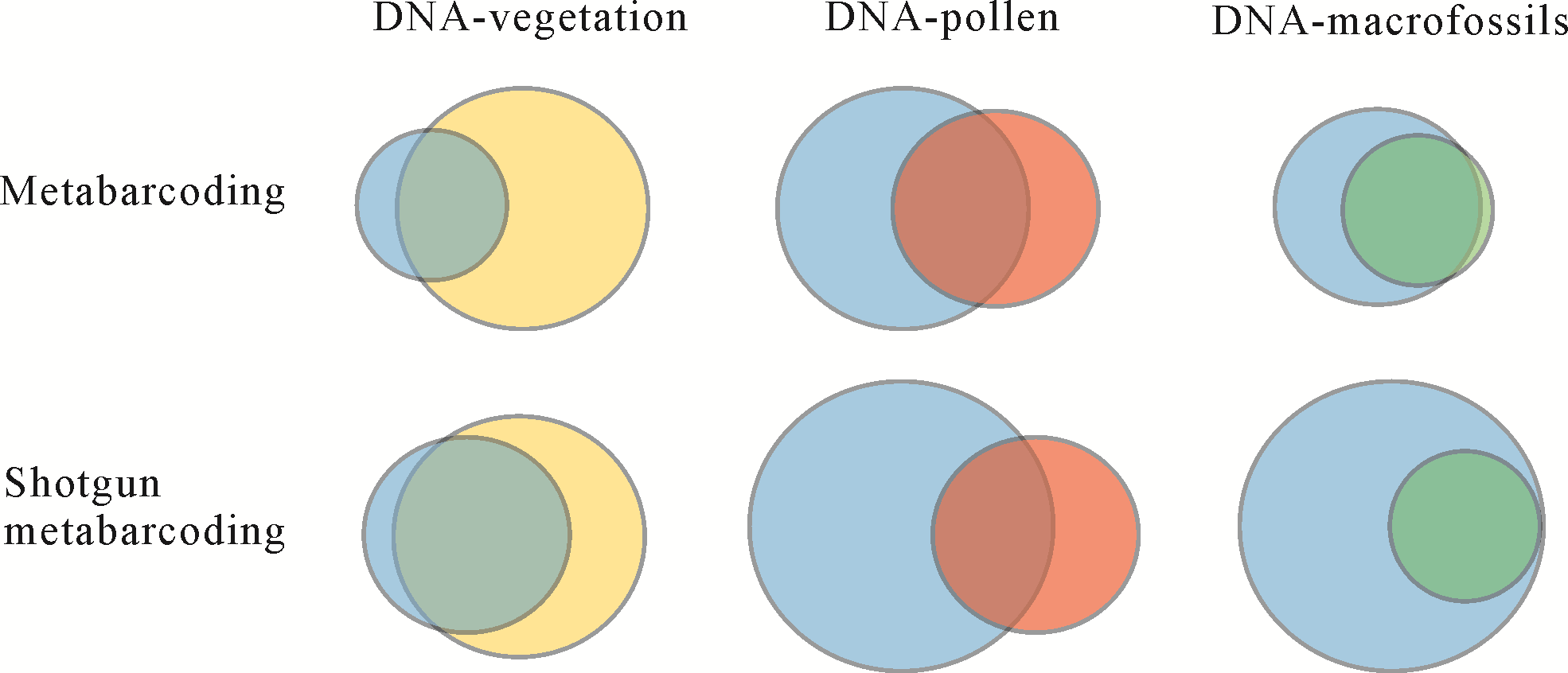
研究表明,湖泊沉积物中的孢粉来自于区域的植物源区,且以风媒植物孢粉为主[29⁃31]。此类孢粉通过气流可实现区域性分布,其传播距离要远高于虫媒植物孢粉。由于孢粉可进行风力传输,其通常记录了较大区域性的植物区系。由于植物残体体积重量较大,不易被风力搬运[32],因此植物大化石来自于本地植物,其种类通常受湖泊周围水生和湿地植物种类影响,陆生植物代表性不足[33]。相关研究证实,沉积物中的植物aDNA与沉积孢粉单体颗粒中的DNA并不完全一致,与植物大化石有着相似的来源[34⁃35],具体关系见图1。大量研究表明[36⁃40],aDNA技术虽不能完全体现植被调查以及孢粉鉴定的数据,但与其有较大范围的重合,对于植物大化石的种类可基本覆盖。从aDNA的两种技术方法来看Shotgun metabarcoding有着更高检测精度,可对更多的种类进行覆盖。

Figure 1. Venn diagrams showing the proportion of plant taxa commonly detected by different proxies (modified from Parducci et al. [10]) (metabarcoding and shotgun metabarcoding of sediment plant aDNA (blue), vegetation survey (light yellow), pollen (red) and macrofossils (green)
-
从保存大环境来讲,aDNA保存程度直接受控于降解速率,最好的保存条件是具备缺氧与寒冷条件的永久性冻土与冰川下,可保存千年之久[41⁃42]。生物反应速率在一定范围内随温度降低而减弱,低温环境是长期保存DNA的条件之一[42]。Willerslev et al.[9]证实了冰芯底部提取到的DNA和氨基酸可用来重建过去的动植物群,并得到存在于冻土中的aDNA年代可达450~800 ka。除此之外,干旱地区,如沙漠,也是高质量aDNA的一个来源[43]。aDNA的保存环境并非严格限制于两极或高海拔寒冷地区。近些年,相关学者在干旱及热带地区也实现了aDNA的提取及分析。Ko et al.[44]研究提取到2.2万年前慈竹坨洞中古大熊猫线粒体基因组,明确了此个体的属与现存大熊猫的祖先分离且并存,其母系祖先分离时间可追溯到中更新世(22.7~14.4万年前)。由此可见,高温并非是aDNA分子保存的障碍[45],水分也是一个重要的限制因子,因为水解反应是DNA分子的必要降解过程[46]。
从有机体自身生物过程来讲,进入环境的死细胞可能很快就会被破坏,它们的DNA会立即被释放到环境中[47],暴露于环境中的胞外DNA分子有3种去向:1)被细菌和真菌核酸酶降解为碎片或完全代谢[48⁃49],酶解作用是胞外DNA降解的最重要机制;2)通过与环境化合物如黏土矿物、有机大分子和其他带电粒子的结合实现保存,结合体一定程度上阻止了DNA在核酸酶作用中被吸收降解[50],除此之外,一些冰冻、缺氧及干旱地区限制了核酸酶的出现,从而减少了核酸分子的降解[51];3)一些基因片段甚至是受损的部分也可以进行自然转化,整合到细菌等遗传物质中[52]。
从DNA分子本身来讲,植物aDNA分子会随着时间而破坏,其自身存在一个衰减周期[53]。目前还未查阅到系统进行植物aDNA分子衰减规律及机理的报道。Allentoft et al.[54]首次测定了骨骼中DNA分子的存在时间,发现242 bp长度的线粒体DNA(mtDNA)具有一个521年半衰期,且低温可以降低其分解速度。如果保存条件良好,绝大多数的科属可以被检测出,DNA读取量甚至可作为科属的生物量[55⁃56]。目前为止,沉积植物aDNA最老记录超过55 ka,而冰芯可将其保存长达450~800 ka[57],孢粉中的aDNA为150 ka[58],更老的保存记录将随着研究深入不断被刷新。基于以上植物aDNA的保存时间,可完全实现通过沉积植物aDNA的信息来重建晚更新世及全新世的植被演替过程。
沉积物中含有许多不同生物体的DNA,包括来自组织和完整细胞的细胞DNA,以及细胞外DNA。对于细胞内的aDNA,不同植物类型及不同种属保存能力是不同的,其主要决定于植物自身特性及沉积环境决定的对于组织降解的差异性。一定程度来讲,具有长时间保存能力的植物组织(如具有较高细胞密度的植物分生组织)可以更好的保存尤其是来自于大量细胞器的DNA,同时,具有一定抗降解能力的植物组织,如种子和果实等[59]均可以得到很好的保存,如较大的枝干及种子(如古莲子等)它的DNA更容易保存在地质体中,反之亦然。目前针对某一种或某几种植物种类或不同科属的保存降解过程、条件及机制方面的研究较为贫乏,不同类型植物还有不同科属植物涉及保存降解各方面的环境阈值并没有一定清楚的厘定[36]。然而,当植物组织细胞破坏,细胞物质释放到沉积物中,其保存降解的过程与前文关于DNA分子保存降解规律一致,并无植物种类和种属差异。Pietramellara et al.[60]认为黏土非常适合保护胞外DNA分子不被降解,因为黏土颗粒具有较大的带电表面积,一旦与胞外DNA分子吸附,会保护其不受核酸酶降解。土壤中的细胞外DNA分子甚至可以被有能力的原核细胞在一个称为自然转化的过程中吸收。虽然这不大可能成为植物DNA的一大来源,但此过程在自然界中客观存在[51]。沉积物中的植物DNA主要以细胞外DNA的形式存在,还是以植物的叶、根冠、细胞、茎或果实等细胞内的形式存在,目前尚不清楚。
-
湖泊是全球气候环境变化的重要信息载体,其连续沉积,高分辨率地保存了区域环境演变及相关气候事件的记录[61]。自然条件下的湖泊沉积物有两个来源:流域侵蚀带来的外源组分(陆生环境);湖泊水体中各种化学生物过程所产生的内生沉淀(水生环境)。因此,湖泊是非常好的沉积环境,随着时间的累积,植物aDNA可以保存在稳定地层和缺氧条件下,是aDNA分子的绝好保存环境[62⁃63]。
由于湖相沉积物受到外界扰乱较小,因此可非常准确的记录周边陆生环境的变化。同样,湖泊环境可以很好地将植物aDNA进行保存。首先,氧含量方面,湖相沉积包含有来自于自身和外来两部分的有机质,微生物对其代谢需要消耗大量的氧气[64],因此创造了水底及沉积物下部的缺氧/低氧环境[58]。这样的缺氧条件基本阻碍了一些需氧生物的生存,将外界生物扰动、沉积体再作用等减到了最小[18],缺氧的环境降低了氧化反应,这对于古生态记录的保存非常关键。温度方面,湖底沉积物始终都处在一个尽可能的低温下:水的密度(ρ)与温度相关,其在4 ℃最大,低于或者高于4 ℃的水团均会呈“漂浮”态,4 ℃左右的低温水团“下沉”底部使其与上部生态系统隔离,这种不同温度的水密度差异特性更加有利于缺氧和稳定的低温条件。湖泊的广泛分布,其水温也取决于其分布地形和湖盆深度[65]。如果湖足够深,不同深度的水会有一个热分层的现象。在夏季,表层水热而底部水冷;在冬季,表水会一直降温到4 ℃后下沉直到整个湖水都混合成4 ℃,然后根据本地气温湖水继续降温到冰冻。基于以上原理,不管湖泊位于低纬还是高纬地区,沉积物总是保持与最冷的水接触进而与生物圈隔离,有利于缺氧环境及温度的稳定,提升了植物aDNA的保存率。由于湖相沉积的低扰动特性,类似于保存于沉积体中的植物大化石、孢粉颗粒等,植物aDNA基本是不存在垂直迁移行为的[15,18],可以认为他们是随着年代忠实保存了环境变迁的记录。然而,再沉积作用以及外界扰动,仍然是相关研究工作中需要注意的可能的“不规则”DNA来源。
1.1 沉积植物aDNA的概念
1.2 沉积植物aDNA的来源
1.3 沉积植物aDNA的保存
1.4 沉积植物aDNA的沉积过程——以湖相沉积环境为例
-
利用Web of Science核心合集进行关键词“Sediment plant ancient DNA”OR“sediment plant aDNA”检索,出版年跨度为1987—2019年,数据库更新时间为2019年10月21日,文献类型共检索到了75条研究型论文(Article),10条综述(Review)及22条会议论文(Proceedings Paper)。其中在8个Web of Science类别占据较大比重,分别为地球科学综合(Geoscience multidisciplinary)(24篇,31.7%)、地球物理(Geography physical)(20篇,26.%)、生态学(ecology)(19篇,24.7%)、生物化学与分子生物学(Biochemistry molecular biology)(12篇,15.6%)、多学科科学(Multidisciplinary sciences)(12篇,15.6%)、进化学(Evolutionary sciences)(11篇,14.3%)。环境科学(Environmental sciences)(10篇,13.0%)及生物学(Biology)(7篇,9.1%)。论文主要来源期刊见表1。从涉及学科及发表期刊来看,沉积植物aDNA研究越来越多被应用到了地理学、环境学、生态学及第四纪地质学等学科,用以解决古气候古生态学中的科学问题。其中高频次引用论文(引用频次> 0,共14篇)集中于在2003—2015年,Molecular Ecology(4),Science(2),Quaterary Science Reviews(2)为主要来源期刊(表2),内容涉及方法探讨与技术应用两大方面。
Table 1.
Major journals publishing articles on sediment plant aDNA 作 者 出版年份 论文题目 来源期刊 被引频次 DOI Davis et al. [66] 2005 Evolutionary responses to changing climate Ecology 403 10.1890/03⁃0788 Thomsen et al. [62] 2015 Environmental DNA ⁃ an emerging tool in conservation for monitoring past and present biodiversity Bioligical Conservation 378 10.1016/j.biocon.2014.11.019 Willerslev et al. [16] 2003 Diverse plant and animal genetic records from Holocene and Pleistocene sediments Science 306 10.1126/science.1084114 Thomsen et al.[67] 2012 Detection of a diverse marine fish fauna using environmental DNA from seawater samples Plos one 281 10.1371/journal.pone.0041732 Parducci et al. [68] 2012 Glacial Survival of Boreal Trees in Northern Scandinavia Science 189 10.1126/science.1216043 Andersen et al. [69] 2012 Meta⁃barcoding of 'dirt' DNA from soil reflects vertebrate biodiversity Molecular Ecology 119 10.1111/j.1365⁃294X.2011.05261.x Pedersen et al.[70] 2015 Ancient and modern environmental DNA Philosophical Transactions of the Royal Society B⁃Biological Sciences 87 10.1098/rstb.2013.0383 Lydolph et al.[71] 2005 Beringian paleoecology inferred from permafrost⁃preserved fungal DNA Applied and Environmental Microbiology 73 10.1128/AEM.71.2.1012⁃1017.2005 Jorgensen et al. [72] 2012 A comparative study of ancient sedimentary DNA, pollen and macrofossils from permafrost sediments of northern Siberia reveals long⁃term vegetational stability Molecular Ecology 72 10.1111/j.1365⁃294X.2011.05287.x Parducci et al. [73] 2013 Molecular⁃and pollen⁃based vegetation analysis in lake sediments from central Scandinavia Molecular Ecology 53 10.1111/mec.12298 Pedersen et al.[34] 2013 A comparative study of ancient environmental DNA to pollen and macrofossils from lake sediments reveals taxonomic overlap and additional plant taxa Quaterary Science Reviews 45 10.1016/j.quascirev.2013.06.006 Bellemain et al. [74] 2013 Fungal palaeodiversity revealed using high⁃throughput metabarcoding of ancient DNA from arctic permafrost Environmental Microbiology 44 10.1111/1462⁃2920.12020 Jorgensen et al. [75] 2012 Islands in the ice: detecting past vegetation on Greenlandic nunataks using historical records and sedimentary ancient DNA Meta⁃barcoding Molecular Ecology 42 10.1111/j.1365⁃294X.2011.05278.x Epp et al. [76] 2015 Lake sediment multi⁃taxon DNA from North Greenland records early post⁃glacial appearance of vascular plants and accurately tracks environmental changes Quaterary Science Reviews 21 10.1016/j.quascirev.2015.03.027 Table 2.
Frequently cited articles on sediment plant aDNA -
aDNA的研究对于生物遗传学、进化学、考古学、古人类学等相关学科的应用相对广泛,而沉积体系中的植物aDNA正在成为一个新的应用于古环境演变研究中的成熟技术[77⁃78]。
沉积植物aDNA的研究始于植物孢粉DNA:德国学者Parducci et al.[68,73,78]首先从瑞典霍尔特贾宁冰湖(Holtjärnen postglacial lake)沉积物中提取到了苏格兰松树和挪威云杉的花粉质体DNA,这些花粉有近10万年的历史,说明植物孢粉DNA在10万年以内可以得到很好的保存。受此启发,Parducci在斯堪的纳维亚采用aDNA技术发现了保存于10.3 ka的本地特有云杉属的DNA,22 ka和177 ka的松属及云杉属的叶绿体DNA,由此证明针叶树可在斯堪的纳维亚半岛上的最后一次冰期间存活,这一结论挑战了当前关于树木生存和扩散是与气候变化相应的主流结论。随后作者在斯堪的纳维亚中部通过两个全新世湖泊样品的采集,发现孢粉与aDNA鉴定科属上存在一定差异,孢粉统计到了46个科属,植物aDNA技术识别到了14个科属,二者综合统计了52个科属。对于古植被重建,运用沉积aDNA技术是一个重要的补充。Parducci作为此技术运用的前驱者,方法与结论同样遭到了质疑:Birks et al. [79]认为,Parducci et al. [68]并没有足够的证据来证实松属和云杉属的冰期生存问题,首先方法上没有严格排除潜在的污染(如取样器,实验设备等),除此之外也没有考虑到栖息地缺乏、冰消温暖期两个物种消失等情况,而且结论与目前植物生态理论相悖。
新技术的开启可以解决传统方法所不能解决的问题。Epp et al.[76]在格陵兰北部湖泊沉积物中采用aDNA手段发现了冰后期维管植物,如岩高兰(Empetrum nigrum)等,并认为基于aDNA重建的植被与预期一致,可准确跟踪环境变化。德国学者Zimmermann et al.[42]在西伯利亚东北部Buor Khaya半岛沉积物中,通过分析沉积aDNA鉴定出了154个科属的植物,孢粉统计出了83个科属,二者共同重建了该区域末次冰期之后的气候,并认为在沉积剖面中,aDNA表现为高分辨率高物种检测丰度的特点,并指出沉积aDNA的种类更多的表现了植物地下生物量的信息。在沉积体中缺乏植物大化石的情况下,采用沉积aDNA和孢粉结合的技术,具有较大优势。Zale et al. [17]在斯堪的纳维亚(Scandinavia)威赫塞尔(Weichselian)后期冰盖附近的冰川瓯穴中采用了微体古生物化石、aDNA、孢粉及硅藻分析,恢复了早期冰上植被系统,认为灌木(柳属、桦木属和杜鹃花科)以及落叶松属等植物均存在于末次冰消期。Bremond et al.[80]提取了非洲西部贝宁希利湖(Lake Sele)湖相沉积DNA,进一步论证了热带地区古NDA存在的可能并发现了22科21属的植物,该结果与孢粉结果重叠性很小,分类对本地植物指示性较强。研究还鉴定到了外来物种地瓜,可见,此技术除了可以较强针对性地重建本地植被还可用于早期农业耕作历史方面的研究。Heinecke et al.[81]通过沉积aDNA技术结合碳氮元素比及同位素(δ 13C, δ 15N)研究了帕米尔高原仿羔皮湖(Lake Karakul)29 ka以来水下植物组成生物量变化,清晰展示了水位变动与古植物多样性的关系。Alsos et al.[20]通过分析斯瓦尔巴特群岛湖泊沉积物,将aDNA技术与植物大化石鉴定对比,aDNA方法额外鉴定出了更多的植物种类:6种维管束植物、2种藻类及12种苔藓类,且每份样品中检测到的种类都要多于大化石,未保存大化石的时期,植被情况同样可识别且变化不大。其研究表明,植物群在古气候变化中具有一定的“韧性”,气温变动在2 ℃之内的情况下植被不会有较大的变动。Clarke et al.[82]报道了瓦兰吉尔半岛(Varanger Peninsula)中部(挪威东北部)基于孢粉及沉积aDNA重建的植被及气候,共统计了101个维管植物和17个苔藓植物科属,其86%的科属依然存在于现在植被中,表明现代灌木—冻原群落及其大部分物种早在10 700年就已经存在,而aDNA结果特别显示了水生植物水芒草(Limosella aquatica)和杜鹃(Rhododendron tomentosum)的持续存在,说明整个全新世早期和中期的气候都要比现在温暖,这是依靠传统孢粉所记录不到的气候信息。可见,高分辨率地挖掘到植物保存的化石与分子记录对于古植被演替及环境重建是非常关键的。随着技术本身的发展强化,其检测精度进一步提高,检测信息也愈加丰富,沉积aDNA手段将不仅仅协助揭示植被的局部信息,还将绝大部分取代孢粉的远源及超/低代表性等弊端,更加全面客观展示本地古植被的演替过程。
2.1 基于Web of Science数据库的基本统计学分析
2.2 沉积植物aDNA的发展及应用
-
古植被重建长久以来依据传统的化石鉴定观察方法:孢粉形态、植物大化石及植硅体的显微观察。DNA分析无疑是一个革命性的替代补充方法,其可提供更为全面及高分辨率的分类信息。HTS 是基于传统桑格测序技术(Sanger sequencing techniques)拓展而来(图2)。aDNA分子被转化为测序库可以在HIS平台(the Illumina HiSeq或Complete Genomics platforms)上并行测序。原则上有3种HTS方法来进行沉积植物aDNA的分析:Metabarcoding,Shotgun metabarcoding及单一孢粉DNA的HTS方法。以下主要介绍应用最为广泛的宏基因条形码技术Metabarcoding及采样实验注意事项。

Figure 2. Approaches to identification of species using plant aDNA (modified from Taberlet et al. [83])
-
metabarcoding即DNA条形码技术,是选用细胞核或细胞器基因组特定基因上、短的DNA序列,进行针对性的扩增和测序来鉴别生物物种的技术。其原理是不同物种在同一DNA条形码区域存在差异,后续通过DNA条形码与参考数据库比对来进行物种的分类鉴别[83⁃84]。metabarcoding的优点是可以同时扩增和鉴别到大量高分辨率的物种分类[85⁃86],得到的植物metabarcoding数据可与常规生态手段获得的其他古环境代用指标协同进行多样性分析。目前较为广泛地应用于各类环境(空气、水体、土壤、粪便等)样品[87⁃88]来鉴定动物、微生物、植物等多样性。
-
metabarcoding技术的关键在于找到一个多物种均存在的基因可变区(Metabarcode),动物方面:最常用的标记基因是18S的V1⁃V2、V4、V7⁃V9变区[89],此变区对无脊椎动物的属、科具有非常好的辨别性,被广泛应用于浮游动物群落的检测中。除此之外,线粒体CO1区也是分析动物多样性的标志基因[90]。植物方面:叶绿体DNA trnL(UAA)内含子P6环区[84,91,92],是目前运用最为广泛的用来鉴别维管束植物分类的基因变区,叶绿体trnL (UAA)内含子引物通用性高、扩增体系稳定及片段大小合适[92],即使是游离和降解严重的土壤DNA 样品也可以扩增出来[82],此基因变区在多种环境DNA样本中得到了应用,如更新世冻土[9,72],晚第四纪湖相沉积[73,93]等。除此还有核基因组的ITS 片段和来自叶绿体基因组的rbcL和matK片段等[94⁃95]。微生物方面:在细菌和古菌分类研究方面,采用小亚基核糖体RNA(SSUr RNA)可对大多数微生物进行分类[95⁃96]。真菌方面,内转录间隔区(Internal transcribed spacer regin,ITS)作为rDNA的非编码区,ITS⁃1和ITS⁃2是最常用于真菌分类的片区[97⁃98]。
-
metabarcoding技术在高通量鉴定物种方面具有很大的潜力,但目前也有一定的局限性。首先是PCR(Polymerase chain reaction,聚合酶链式反应)技术方面的问题:PCR扩增过程会来带一定的误差[99],这种误差的引起主要源于aDNA模板可能存在不同程度的降解,PCR扩增时替换、插入和缺失以及测序过程中产生的误差[83]。其次是metabarcode的选取上,一些研究表明其采用的引物扩增后得到的OTUs(Operational taxonomic units)并没有预期的高[100],可以根据研究目的来添加特异性种的引物扩增进行更高分辨率的分析,如采用添加禾本科上下引物:ITS1⁃F/ITS1Poa⁃R 及菊科:ITS1⁃F/TS1Ast⁃R[100]来解决,通用和物种特有引物的结合使用可最大限度地提高多样性的分辨率。由于不同的引物对于一些种类的扩增偏向不同,这可能导致一些种类被过多的扩增而一些较少扩增甚至不被扩增,因此不同物种分类并非有同样的扩增效率[101]。
最近发展的shotgun metabarcoding是metabarco⁃ ding的替代技术,被认为是绕过DNA条码扩增偏差的重要手段[53,102]。Shotgun metabarcoding会在将来几年成为一个重要的工具,因为它可更高分辨率地鉴定生物多样性,并发现种群结构的细微差异。shotgun metabarcoding结合NGS平台,其可避免PCR而直接采用aDNA模板进行测序,其可读取数十亿的序列。除此之外,metabarcode选择及组合也是决定分类准确性的关键。
其另一个局限性表现在分类与MOTUs(molecu lar operational taxonomic units)的对应上,对大多数MOTUs来说,关键在于缺乏全面的、分类可靠的metabarcoding数据库[103⁃105]。加强metabarcoding在更大时空尺度上对生物多样性的监测,需要增加物种标准条形码的识别,特别是在物种高度多样化的热带地区[97,103]以及一些稀有物种的信息。metabarcoding技术在古环境重建与现代环境生物物种检测工作中发挥着举足轻重的作用。
-
植物aDNA的沉积样芯的收集与常规古生态研究中的钻取设备一致,但采样管需提前进行杀菌消毒处理,以杜绝现代环境残留物质对样品的污染。野外样品需要通过密封管将样品运到实验室。样品处理及分析的各个阶段都应采取特别的预防措施。可能的污染来自于三个方面:设备本身、样品之间交叉污染以及实验药品。因此,每一个样品的分样都需要使用乙醇及短波紫外线(UVC)进行杀菌[105]。消毒设备,保证无菌的工具是首要的,实验者在实验室的整个实验过程需确保穿着隔离实验服、口罩与手套要佩戴齐全。
实验样品的选取需是样品芯最中心的部分。取样时候可以将DNA示踪剂用到取芯设备上,这样就可以检测来自外层的DNA分子是否进入沉积物内部[34],或者在分样时准备无DNA的超纯水暴露在空气中来对是否受到污染做对照。防止外源DNA污染需要在整个实验过程严格把控。
-
一套成熟的DNA条形码系统(barcoding system)需符合以下原则:1)该系统目标DNA区域在同一物种中高度保守,在不同物种间差异较大;2)该系统目标DNA区域应当是标准统一化的,即对不同物种分类时,均可将此DNA区域作为分类依据;3)该系统目标DNA区域应包含足够的物种遗传进化信息,以便准确的将物种划分归类(科、属等);4)该系统目标DNA区域应具有高度保守的启动位点。除此之外,快速、高效的基因扩增和测序手段也是该系统重要组成部分[85]。
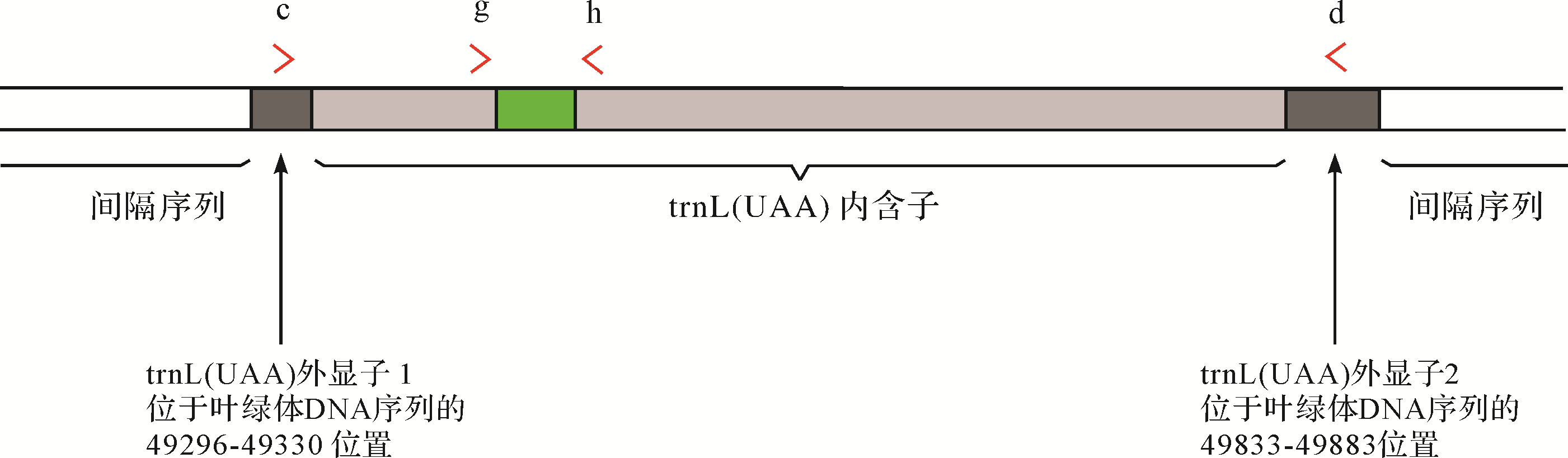
目前在植物分类上,最理想的同时也是应用最为广泛的DNA条形码是叶绿体基因的trnL(UAA)区[34,83,106],该区是叶绿体基因中唯一的内含子区域[107],具有高度保守的二级结构[108](图3)。用于该区域扩增的引物主要有两对,分别是g、h和c,d(表3)。其中g、h主要用于扩增叶绿体trnL(UAA)P6环区。

Figure 3. Tobacco trnL(UAA) and primers action fragment (modified from Pierre et al. [85] The P6 loop amplified with primer g and h is indicated in green)
名称 编号 序列(5’⁃3’) c A49235 CGAAATCGGTAGACGCTACG d B49863 GGGGATAGAGGGACTTGAAC g A49425 GGGCAATCCTGAGCCAA h B49466 CCATTGAGTCTCTGCACCTATC Table 3.
Sequences of the two universal primer pairs amplifying the trnL(UAA) intron -
以Alsos et al.[109]为例:
-
首先将冷冻后的沉积物样品置于冰箱中解冻24~48 h,之后称取4~10 g用于DNA提取。采用A PowerMax Soil DNA Isolation kit (MO BIO Laboratories, Carlsbad, CA, USA)试剂盒,依据试剂盒的提取说明进行DNA的提取。
使用引物“g”、“h”来对叶绿体trnL(UAA)内含子P6环区[86]进行PCR扩增。引物包含长8 bp的5’端侧翼序列,保证混合样品的并行测序[110⁃111]。
PCR扩增采用50 μL体系:DNA样本5 μL,15 mM Tris⁃HCl,50 mM KCl,2.5 Mm MgCl2,0.2 mM each Dntp,0.2 μM each primer和8 μg血清蛋白。所有PCR样品(DNA样品和对照组)均置于PCR板上。PCR程序:94 ℃预变性10 min;94 ℃变性30 s,50 ℃退火30 s,72℃延伸1 min,共45个循环;最后72 ℃再延伸7 min。反应结束后,将PCR产物混合在一起,使用MinElute Purification kit (Qiagen GmbH, Hilden, Germany)试剂盒对产物进行纯化。使用Illumina 2500 HiSeq测序平台TruSeq SBS Kit v3(FASTERIS SA, Swizerland)对产物进行测序[20]。
-
测序结果使用OBITools程序完成初始筛选[112],之后通过对比本地分类参考库(包含近千种代表维管束及苔藓植物分类[57,86]的近千个DNA序列)使用ecotag程序进一步将序列归类分配到分类单元[112],随后二次对比欧洲核酸序列数据库(EMBL)。达到100%匹配的序列会被保留。最后通过BLAST (Basic Local Alignment Search Tool)(
http: //www.ncbi.nlm.nih.gov/blast/ )来检测序列匹配的潜在错误。
3.1 Metabarcoding(DNA条形码技术)
3.1.1 定义
3.1.2 生物常用的metabarcoding基因区
3.1.3 局限性
3.2 沉积植物aDNA的实验方法
3.2.1 实验准备
3.2.2 沉积植物aDNA的metabarcode及常用引物
3.2.3 提取方法
(1) DNA提取和扩增
(2) DNA序列分析与筛选
-
近些年飞速发展的生物分子技术为第四纪环境和考古学研究提供了一个较新的技术支撑,这是一门工具科学。除了古环境(古植被)重建方面,在大型物种灭绝、人类—环境相互作用、生物分类学研究及DNA损伤等研究也有较大应用。然而,新技术的应用并非完美,其对古植被重建的精度方面也受制于较多的因素:首先是技术本身方面,如PCR扩增及测序阶段导致的错误;其次是核酸库的成熟度上;最后是DNA模板质量的优劣,即模板量是否充足及降解,模板的优劣受埋藏条件、物种差异、提取试剂及操作方法影响。因此,从来源上,埋藏条件上,提取扩增及测序技术上,到最后比对上,甚至实验者的实验技术及操作经验等各个环节均会对古植被重建的精度/准确度上产生影响。
沉积植物aDNA已经成为古植被与传统古生态学分析相结合的工具。相对于其他方法,它与植物大化石均侧重于本地植被重建,且在识别“化石沉默”中起到关键作用,DNA分子可更为精确地记录植物组合种群起源、遗传分化及扩散。当然,一些研究的结论也引起了一些学者的质疑,如提取DNA的可信性以及其来源等,都会在未来的研究中得到解答。针对后期的研究,试提出几点研究方向:
沉积植物aDNA生态学方面研究:1)全面开展不同生境下湖泊沉积的赋存状况与环境因子的相关性研究,明确其保存的多种环境条件阈值;2)植物aDNA环境行为学研究,通过环境因子胁迫,如室内模拟不同温度梯度、不同压力、不同保存介质(土壤、水体)下对不同植物种类化石aDNA分子提取量的差异性影响,明确其在环境中的损伤机制与降解周期;3)沉积体系中植物aDNA来源研究,进一步明确其与其他古植被代用指标的重叠性与补充性关系,如在不同植被类型研究区通过植被调查、不同层次土壤DNA提取、孢粉及植物残体鉴定来验证几种方法对植被信息的反应程度及之间的重叠包含关系。
植物aDNA技术方面研究:1)技术应用的科学性论证方面,通过现代生态学过程进行验证;2)实验设计的优化、引物的选择,PCR体系优化、物种条形码的扩充及复合数据库的完善等,即目前还没有一套标准适用于沉积植物aDNA的实验室操作规范,metabarcode依据研究目的的多元化选择,与其匹配引物的设计,最优PCR体系的验证实验、比对数据库的完善等,均需要投入大量的研究。3)植物aDNA应用方面研究:1)扩大研究区域尺度,应用此技术协同传统古生态学方法完善植被重建的物种基础数据,进一步完善建立地方数据库,目前涉及该方法应用的国外报道较多,我国还未大规模开展。2)通过古植被恢复及古植物单体基因测序分析植物遗传分化规律,可对一些第四纪冰川孑遗植物的遗传谱系进行系统研究。

 DownLoad:
DownLoad: