HTML
-
白云岩成因一直是地质学界研究的一个热点,尽管前人提出了多种成因机理[1-9],但许多机理没有过硬的证据,因此其成因迄今也没有完全弄清楚。随着深层勘探的不断深入,埋藏深、时代老的微生物碳酸盐岩成为主要的目的层[10],因此,解决白云岩成因问题十分有必要。
曾经困惑沉积学家们的一个重要问题是:现代沉积环境中发现了不少白云岩,但常温下在实验室内却无法人工合成,因此很多人认为不存在原生白云岩。而近些年的研究发现,白云石借助微生物的作用在常温下是可以沉淀出来的,白云石的形成可出现在适合微生物生长,有足够Ca2+、Mg2+和
以往研究发现,能够沉淀白云石的微生物有多种,而且有的是在缺氧条件下形成,有的是在富氧条件下形成[12-37]。有些学者曾对其中一些微生物白云岩形成机理进行过简要介绍[38-43]。但如何识别古代的微生物白云岩?如何识别不同成因的微生物白云岩?关于微生物白云岩的形成机理,还有哪些疑问和需要解决的问题?前人的研究尚未回答这些问题。本文将以已在自然界中发现且资料较丰富的四类微生物白云岩为例对这些问题进行探讨。此外,本文对每种机理和实例进行了较详细的介绍,以便读者能快速获得一些关键信息。
-
本文介绍的4类微生物白云岩形成机理包括:硫酸盐还原菌白云岩形成机理、甲烷生成菌白云岩形成机理、甲烷氧化菌白云岩形成机理、喜氧喜盐菌白云岩形成机理。
-
Vasconcelos et al.[13]在巴西东海岸的Lagoa Vermelha潟湖缺氧还原性黑色富含有机质沉积物中发现了白云石。他们用采自这个潟湖的沉积物中的硫酸盐还原细菌Desulforvibrio,模拟潟湖缺氧环境条件,在实验室厌氧条件下沉淀出了有序性较差的富钙铁白云石。于是他们提出,硫酸盐还原细菌能够通过生物化学调节克服白云石形成的障碍,在常温常压下沉淀出白云石。
目前关于硫酸盐还原菌沉淀白云石机理的解释是:
硫酸盐还原细菌利用
这种反应降低了水中
-
硫酸盐还原菌白云岩的现代实例是巴西Lagoa Vermelha潟湖中的现代白云岩[14]。该潟湖位于巴西东海岸里约热内卢以东90 km处,面积约2.4 km2,最大水深1.7 m,周围的微气候为半干旱,潟湖边缘发育藻席。潟湖与大西洋之间为沙丘组成的宽400 m的障壁岛。根据14C定年,潟湖形成于3 600年前。
潟湖的湖水盐度随干、湿季节波动较大,由盐水(hypersaline)到半咸水(brackish);月平均水温为23 ℃~32 ℃,pH值为8~8.5;δ 18OSMOW为1.50‰~3.82‰,多为2‰~3‰;Mg/Ca为1.66~3.13,多为2.5~3。
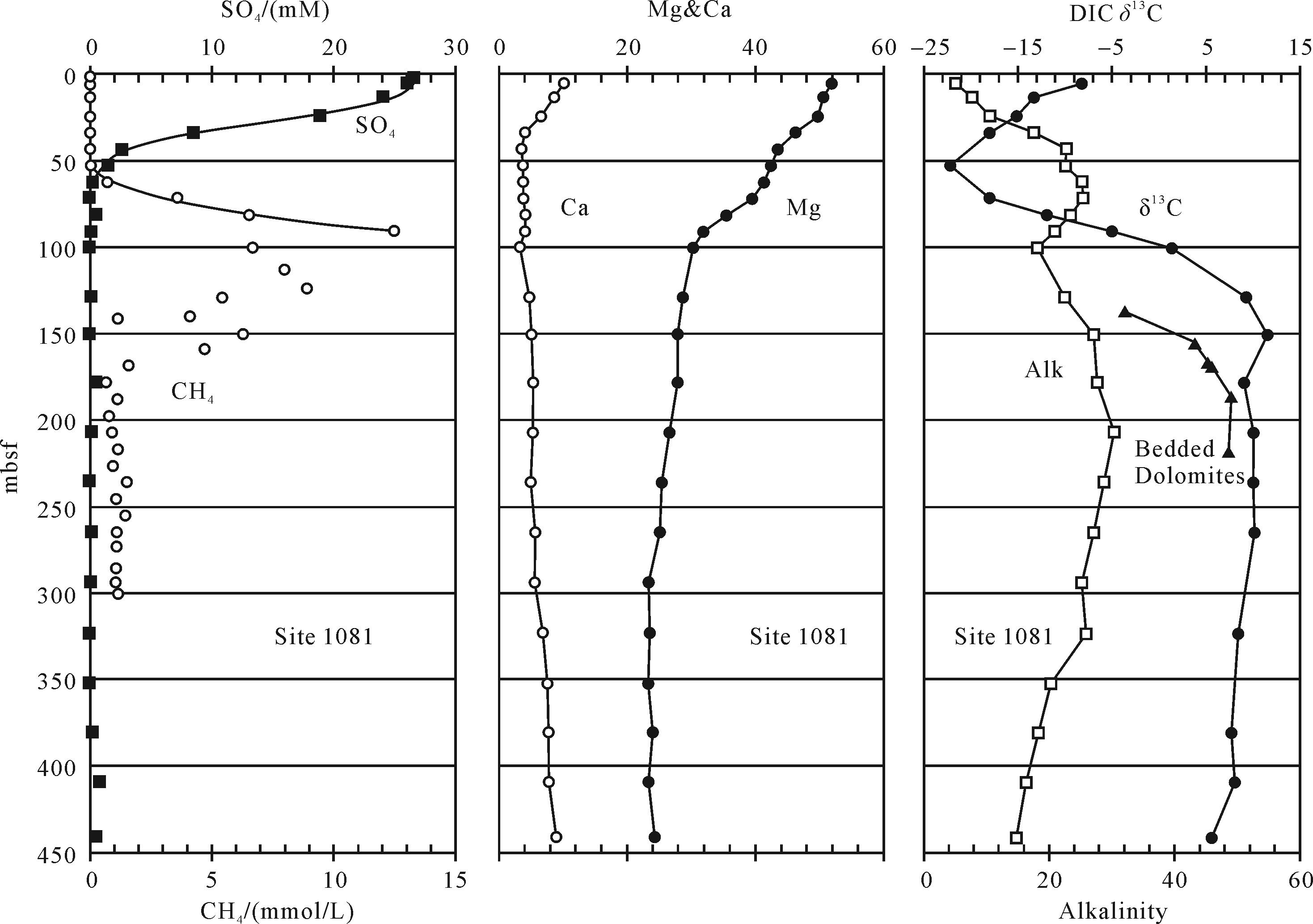
潟湖底部沉积物表面覆盖了一层10 cm厚的黑色淤泥,水底缺氧。淤泥主要是碳酸盐泥、有机质、细菌与水的混合物。碳酸盐泥由方解石、高镁方解石和富钙白云石组成。潟湖中央一段1.7 m的取心中(图1),上部1.2 m为碳酸盐沉积物,由含有白云石的球粒层与纹理层交替叠置组成,为潟湖沉积。

Figure 1. Histogram of Lagoa Vermelha lagoon core and corresponding geochemical characteristics[14]
碳酸盐沉积物中有机碳含量普遍高,为2%~18%(图1)。黑色淤泥和表层沉积物中有机碳含量约10%。碳酸盐全样的δ 13CPDB为-2‰~-10‰(图1);δ 18OPDB为1.5‰~5.5‰。δ 18OPDB越高,δ 13CPDB就越低。
岩心中见白云石结核富集层,分布在深度为12~15 cm和72~95 cm之间(图1)。上部的白云石结核为弱固结,白云石含39~42 mol/mol的MgCO3。下部则致密,白云石含量高达98%,白云石含48 mol/mol的MgCO3。这反映白云石随着时间推移变得更接近化学计量,并且有序度增加,晶形也变好。下部白云石结核的δ 13CPDB为-8‰~-10‰,数值很低,反映有机碳的混入。该层上、下沉积物中有机质的δ 13CPDB为-19‰~-21‰,平均-20‰,与温暖海水植物的一致[12],而该层中无定形有机质的δ 13CPDB为-23‰,较低,反映有硫酸盐还原细菌的参与[14]。
Vasconcelos et al.[14]认为该潟湖白云石很低的δ 13C和球状、哑铃状形态是微生物成因的证据。较深处白云石结核层中的白云石有序度高,是“老化”所致;白云石呈较明显的菱形,则认为是后期围绕球形微生物白云石核又发生无机沉淀导致的。他们用稀盐酸部分溶解后,发现露出了包裹在菱形白云石晶体内的球形白云石晶核,就证明了这一点。
-
海(湖)底沉积物从上到下可分2个带:富氧带和缺氧带。富氧带/缺氧带界面可以位于沉积物表面之下,但也可等于或高于沉积物表面(此时富氧带缺失),这取决于水体的循环程度和沉积物中有机质含量。有机质含量越高,该界面越浅。
缺氧带自上而下可进一步划分出硫酸盐还原带(甲烷氧化带)和甲烷生成带。
-
硫酸盐还原带内,硫酸盐还原细菌以
CH4+
这个反应式的意义在于:即使沉积物(如砂质沉积物)缺乏有机质,只要孔隙水中含有
甲烷氧化菌白云岩形成机理与硫酸盐还原菌白云岩的基本一致,都是通过降低水中

Figure 2. Geochemical characteristics of pore water profiles at 1081 points of the Oceanic Drilling Program [15]
-
甲烷生成带内,甲烷含量很高。沉积物中的有机质通过厌氧微生物分解为甲烷和CO2:
2CH2O→CH4+CO2
这种作用又称甲烷生成作用,只能在缺氧、缺乏硫酸盐的孔隙水中进行。当硫酸盐经过甲烷氧化带而耗尽后,甲烷生成作用便开始了,因此该带位于甲烷氧化带之下。甲烷生成菌白云岩形成机理与甲烷氧化菌白云岩形成机理相似,不同的是
-
下面简要介绍甲烷氧化菌白云岩和甲烷生成菌白云岩的现代实例。
-
在邻近非洲纳米比亚西海岸的深海盆地中,多个钻孔的资料表明,海底沉积物上部为硫酸盐还原带(甲烷氧化带),其下为甲烷生成带(图2)。硫酸盐还原带厚度为6~50 m。当有机质含量很高、沉积速度很慢时,硫酸盐还原带厚度可仅为几厘米[27],甲烷生成带的厚度为十几米到100 m左右[15]。
自上而下,沉积物孔隙水的
在硫酸盐还原带(甲烷氧化带)内,自上而下
在甲烷生成带,CH4含量先升高又降低;
在钻孔中发现了数层白云岩,其现今埋藏深度从几十米到400 m左右,目前均位于甲烷生成带内。其δ 13C为-5‰~7‰(PDB)[15]。根据δ 13C判断,δ 13C为-5‰的白云岩应是在甲烷氧化带内形成的,后来埋藏到了甲烷生成带;δ 13C为4‰~7‰的白云岩则形成于甲烷生成带。
-
日本琉球群岛南部Kuroshima海底平顶山上现代沉积物中分布着一些白云岩烟囱[22]。白云岩烟囱直径5~30 cm,高多为30 cm左右,是由白云石和方解石胶结砂、泥而形成的。烟囱中白云石含量为79.2%~83.2%(重量百分比)。白云石晶体微小,多不到1 μm。其δ 13C为-43.6‰~-17.0‰,δ 18O为5.66‰~7.92‰(PDB)。Takeuchi et al. [23]认为,烟囱中的白云岩是由从深部冒上来的甲烷被微生物氧化形成的甲烷氧化菌白云岩。
在丹麦北端距岸10 km处水深10~12 m的砂质海底上分布着一些白云石胶结的砂质柱[23]。砂质柱直径可达1 m,高出海底可达4 m,应具有较深的“根”,因为这些柱子能抗风浪。作为砂质柱胶结物的白云石为0.01~0.03 mm的泥晶,菱面体状,富钙[(Mg47, Ca53)CO3],Sr含量约1 000×10-6,Fe和Mn含量变化大,平均值分别为1 870×10-6和320×10-6。其δ 13C为-44‰~-40‰(PDB),δ 18O为3.7‰~3.9‰(PDB)。Jørgensen[24]推测砂质柱中的白云石是由向上泄露的甲烷被厌氧或需氧微生物氧化形成的,其极低的δ 13C 反映了这一点。甲烷来自下方浅埋藏的气藏。
此外,在世界很多其他海域,海底也都发现了碳酸盐烟囱,如俄勒冈陆架边缘[24]、北海[26]、南新西兰[27]及南中国海[28]等,多与甲烷泄露有关。
Roberts et al. [29]通过野外实验和和室内模拟实验表明,在缺氧的淡水环境中,只要有Ca2+、Mg2+等离子来源,生甲烷菌也可以沉淀出白云石,而且无需高的Mg/Ca比值,在1左右即可。野外实验是针对美国明尼苏达州Bemidji附近被石油污染的、底板为玄武岩的蓄水层进行的。蓄水层处于缺氧状态,地下水对白云石饱和或略过饱和。在污染带活动的微生物主要是铁还原菌,其次是生甲烷菌。
Roberts et al. [29]在蓄水层中钻一口井,将干净的玄武岩样品放入钻井中(距地面7 m),浸泡在地下水中。3个月后,将样品取出并观察,发现玄武岩样品表面生长有微生物,围绕微生物有铁白云石沉淀出来。他们推测生甲烷菌通过消耗水中的CO2使白云石处于过饱和并沉淀出来,所需的Mg、Ca是由微生物腐蚀玄武岩的表面而释放出来的。为了证明这一观点,他们做了模拟室内实验:将玄武岩和采自蓄水层的水、少量石油以及铁还原菌和生甲烷菌放入缺氧的密闭容器内,在25 °C、无阳光的暗室内放置了8个月。之后,取出样品进行观察,发现围绕微生物细胞和玄武岩表面上沉淀出了方解石和白云石,大小为0.02~5 μm,其中白云石约占30%,不含铁,可能是Fe2+含量过低所致。白云石的δ 13C为-0.03‰,δ 18O为-3.39‰;方解石的δ 13C为-0.83‰,δ 18O为-6.53‰。
他们的研究表明,在微生物参与下,白云石的形成既不需要对白云石高度过饱和的咸化水也不需要高的镁钙比。白云石的沉淀可能是由于微生物发生矿化作用时,细菌的胞外聚合物(EPS)促进了白云石的成核过程[44-46]。细菌所分泌的EPS具有从周围的水中集中并绑定钙、镁、碳酸盐、二氧化硅等离子的能力[47]。当电子传递发生时,一些细菌的外膜所能络合的阳离子主要就是Mg2+和Ca2+[48],从而为白云石的沉积创造了有利的微环境。因此微生物的媒介作用不仅体现在调节介质碱度、营造利于白云石沉淀的化学条件方面,更重要的是它们还能为白云石提供初始成核位置,促使白云石晶体发生沉淀[17]。
-
多种微生物,包括蓝细菌(即蓝绿藻)和通过分解含氮有机质产生氨气的细菌,可以在有氧条件下诱发碳酸盐沉淀[30],包括白云石[31]。
Sánchez-Román et al. [32]通过室内实验表明,常温常压下,一些喜氧喜盐菌可以在氧化条件下沉淀出含镁的白云石、高镁方解石、水菱镁矿、碳酸钙镁石(huntite)和鸟粪石。他们使用了5个喜氧喜盐菌属下的21种细菌,其中包括盐单胞菌Halomonas属的16个种,海单胞菌Marinomonas属的2个种,海杆菌Marinobacter属的1个种,色盐杆菌Chromohalobacter属的一个种,以及需盐芽孢杆菌Salibacillus属的1个种。
在30 ℃下,将样品在大气中放置了1个月,其间进行多次观察,然后将沉淀物分离出来进行分析化验。他们的研究表明,对于不同的样品,鸟粪石和碳酸盐矿物沉淀需要的时间不同,短的为4天,长的为22天。
他们的实验表明:
(1) 微生物作用普遍使pH值升高。所有含有微生物的样品在1个月后,pH值均升高至9左右。导致pH值升高的原因是微生物的新陈代谢作用释放出了NH3。
(2) 微生物作用诱发了沉淀。所用的介质对碳酸盐矿物都是不饱和的,但却沉淀出了碳酸盐矿物,这是因为微生物的新陈代谢作用改变了其周围的微环境,使pH值升高,释放的CO2则使
(3) 碳酸盐矿物晶体数量和大小随放置时间增长而变多、变大,晶体大小为几微米到几十微米,呈球状或哑铃状。
(4) 介质中Ca2+和Mg2+的浓度影响矿物的类型和沉淀的先后顺序。在Ca2+浓度最低(0.057 M)的样品中,鸟粪石最先沉淀,之后是水菱镁矿、碳酸钙镁石、高镁方解石和白云石;在Ca2+浓度为0.011 mol/L的样品中,水菱镁矿最先沉淀,之后是高镁方解石、白云石和鸟粪石。在Ca2+浓度为0.017~0.022 mol/L的样品中,白云石最先沉淀,之后是水菱镁矿和鸟粪石。
(5) 介质中Ca2+含量越高,白云石和高镁方解石中的Ca含量就越高。
(6) 碳酸盐矿物的δ 13C为-26.8到-18.2‰,反映为有机碳源。其中,白云石的δ 13C为-26.8到-20.0‰。随着富镁碳酸盐矿物中Ca含量增加,δ 13C有变小的趋势,而Mg增加则会使δ 13C变高。这说明Ca有利于12C进入碳酸盐矿物晶格,而Mg则有利于13C进入晶格。因此,白云石的δ 13C一般比与其共生的方解石的δ 13C高。
-
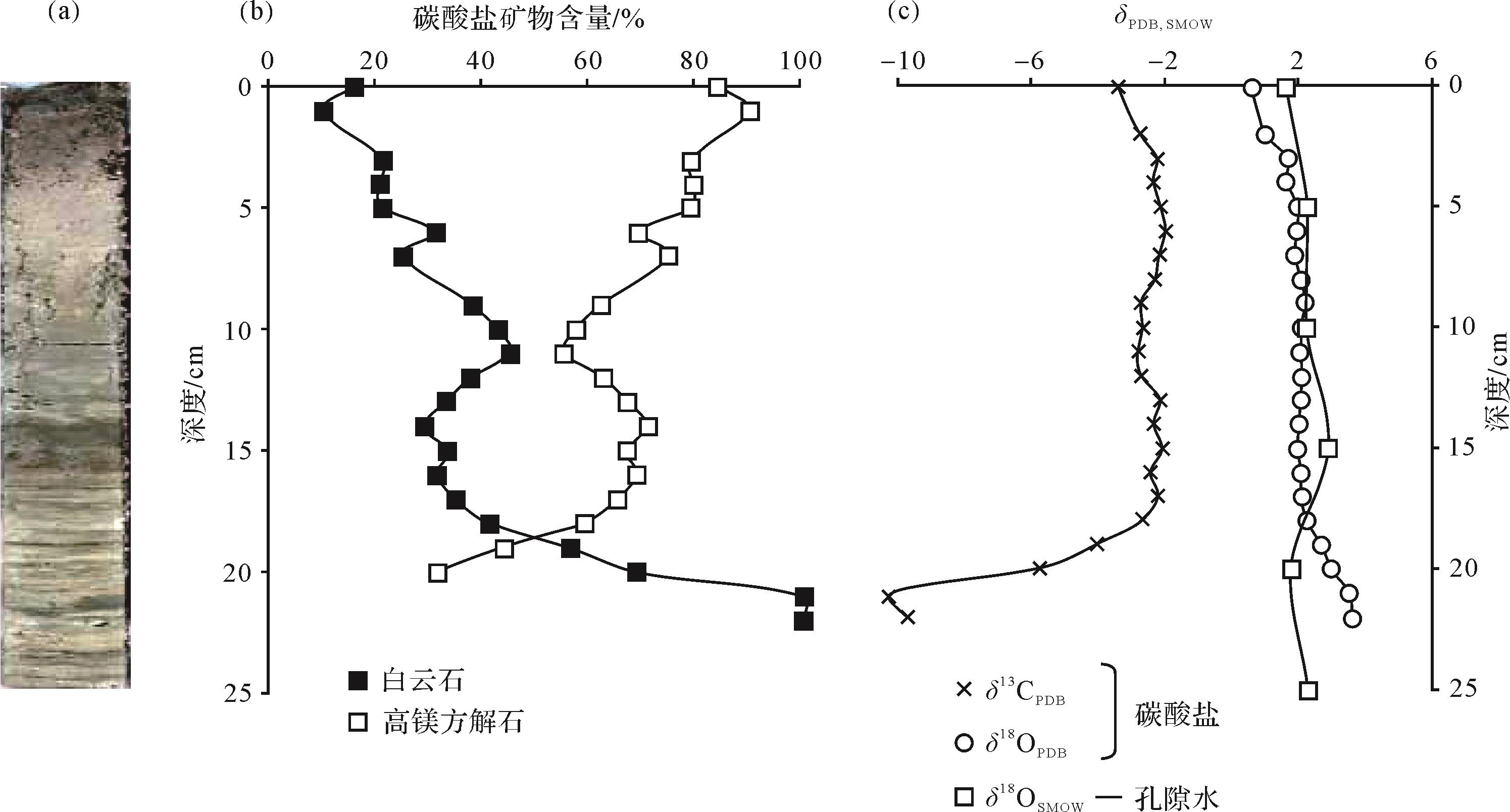
喜氧喜盐菌形成白云石的一个现代实例是巴西Brejo do Espinho潟湖中的现代白云石[32]。该潟湖位于前述Lagoa Vermelha潟湖东侧,也是咸化潟湖,盐度随干、湿季节变化。其水深<0.5 m,Mg/Ca比平均约为5。湖底发育较厚的多种颜色的藻席,并含氧。
在潟湖中部取了25 cm的沉积物(主要由碳酸盐泥组成)岩心,其上部10 cm纹理不清晰,下部15 cm纹理发育(图3)。矿物成分主要为高镁方解石和白云石,还有少量方解石。自上而下,高镁方解石含量降低,白云石含量升高,在21 cm处全部变为白云石,说明高镁方解石转化成了白云石。沉积物中有机质含量向下逐渐降低,在表层有机碳含量为12.5%,而到了纯白云岩层,则降为2.5%,说明在白云石形成的过程中消耗了有机质。碳酸盐沉积物的δ 13CPDB从表层到18 cm深处变化不大,约为-2‰,但白云岩层的δ 13CPDB却快速降低至-10‰,反映有机碳的大量加入。δ 18O则一直较稳定(图3)。

Figure 3. Brejo do Espinho core bar chart and geochemical characteristics[33]
研究者利用从潟湖沉积物中提取出来的两种细菌(Virgibacillus marismortui和Marinobacter sp.),在实验室内模拟潟湖沉积的成岩环境,在有氧条件下沉淀出了白云石,说明潟湖沉积物中的白云石应是这些细菌形成的。
1.1 硫酸盐还原菌白云岩形成机理及现代实例
1.1.1 硫酸盐还原菌白云岩形成机理
1.1.2 现代实例
1.2 甲烷氧化菌和甲烷生成菌白云岩形成机理及现代实例
1.2.1 甲烷氧化菌白云岩形成机理
1.2.2 甲烷生成菌白云岩形成机理
1.2.3 现代实例
(1) 非洲西海岸地区的甲烷氧化菌白云岩和甲烷生成菌白云岩
(2) 其他地区的甲烷氧化菌白云岩和甲烷生成菌白云岩
1.3 喜氧喜盐菌白云岩形成机理及现代实例
1.3.1 喜氧喜盐菌白云岩形成机理
1.3.2 现代实例
-
如何识别古代微生物白云岩?如何识别古代不同成因的微生物白云岩?根据各类微生物白云岩的形成机理及其现代实例,笔者从晶粒大小、形态、碳氧同位素、共生组合、晶体结构等方面对各类微生物白云岩的识别标志进行了讨论和总结。
-
无论是从模拟实验还是现代沉积来看,各类微生物白云岩晶粒普遍都很细,为泥晶级,大小在5 μm以下。即使埋藏后可发生重结晶,晶粒大小也不超过粉晶级。因此,要定为微生物白云岩,砂糖状白云岩则不在考虑之列。
由于上述4种成因的微生物白云岩晶粒相近,因此不能根据晶粒大小区分是哪种成因的微生物白云岩。
-
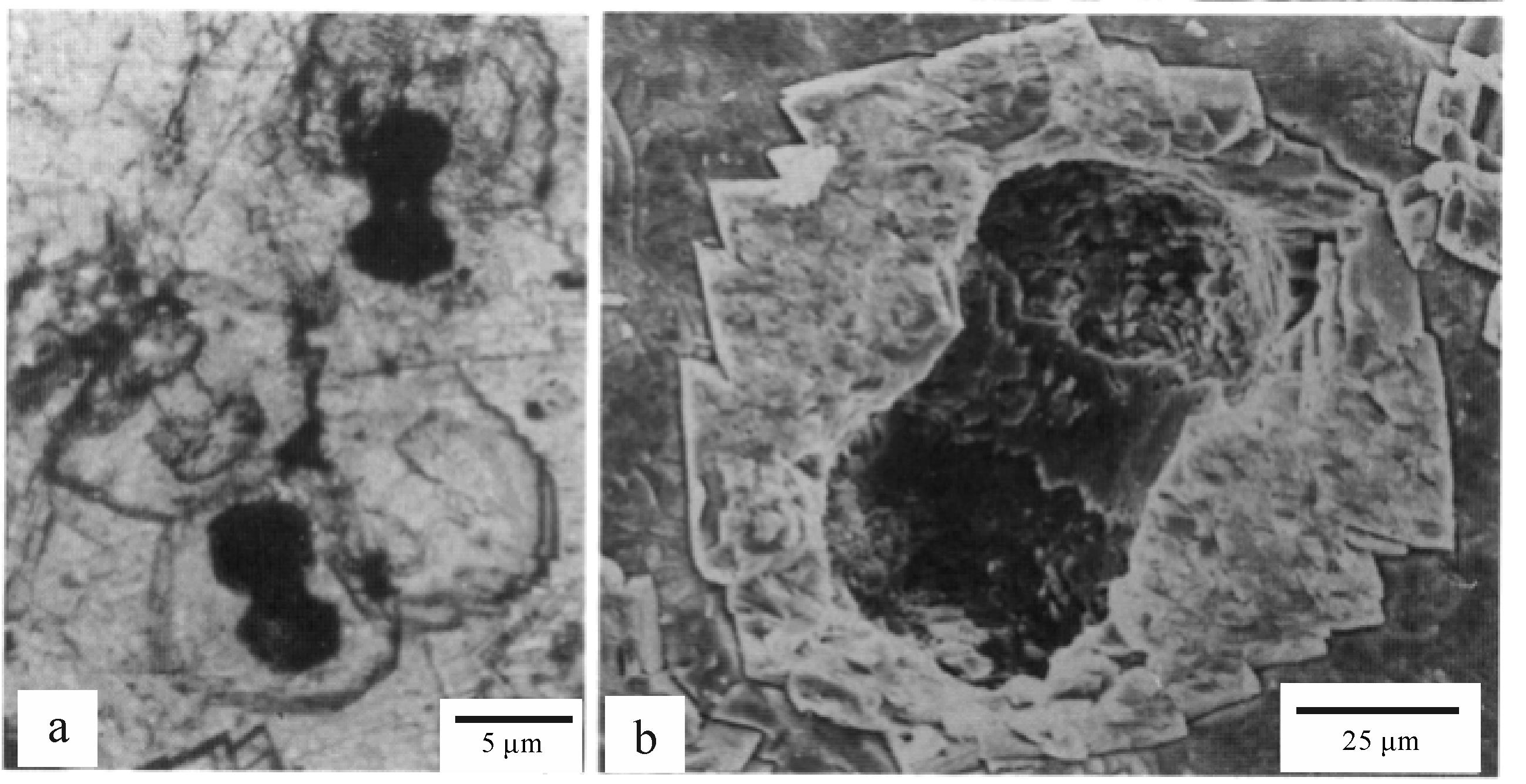
微生物成因的白云石有特殊形态。不少研究者认为,微生物成因的碳酸盐矿物具有特有的球状或哑铃状形态,与无机成因的不同[34-35]。因白云石常围绕微生物沉淀,因此,白云石或其集合体呈球状或哑铃状,其核心为球状或哑铃状微生物(图4)[36]。即使后期经过“老化”晶体呈菱面体,用稀盐酸部分溶解后,仍有可能见到球状或哑铃状白云石,切片中仍有可能见到球状或哑铃状核心。这是识别微生物成因的白云石最过硬的证据,也是研究中需要极力寻找的证据。不过,不是所有微生物成因的白云石内部都具有这种球状或哑铃状构造,因为白云石可由微生物胞外聚合物等诱发沉淀[41]。

Figure 4. Microbial morphology retained within the dolomite[36]
由于形成白云石的各类细菌的形态相似,因此无法根据形态区分不同成因的微生物白云岩。
-
由于有机质在微生物白云岩的形成中起重要作用,因此微生物白云岩的碳氧同位素独具特色,而且不同成因的微生物白云岩在碳氧同位素方面有显著差异,可据此识别微生物白云岩和区分不同成因的微生物白云岩。根据现代实例和模拟实验,本文总结了各类微生物白云岩的碳氧同位素组成。
硫酸盐还原菌白云岩:δ 13C介于-5‰~-10‰(PDB);δ 18O为2‰~5‰(PDB)。
甲烷氧化微生物白云岩:δ 13C通常在-5‰以下,可达-20‰(PDB)[37]。该类白云岩的δ13C与硫酸盐还原微生物白云岩的δ 13C有重叠,但如果低于-10‰(PDB),则应为甲烷氧化微生物白云岩。
甲烷生成微生物白云岩:δ 13C普遍为正值,且较高,这是与其他微生物白云岩显著不同的,也是其主要识别标志。其δ 13C介于0.4‰~13.8‰(PDB);δ 18O为-6.7‰~5.6‰(PDB)[36]。
喜氧喜盐微生物白云岩:δ 13C为-10‰(PDB)左右。δ 18O则一直较稳定,为2‰~3‰(PDB)。
由上述可见,δ 13CPDB在识别微生物白云岩和不同成因的微生物白云岩方面十分有用。
与无机成因的泥粉晶白云岩(蒸发泵白云岩和原生无机化学沉淀成因的白云岩)相比,硫酸盐还原、甲烷氧化和喜氧喜盐微生物白云岩的δ 13CPDB普遍很低。现代无机成因的泥粉晶白云岩的δ13CPDB都是正值,通常在0~3‰之间。因此,如果古代泥粉晶白云岩的δ 13CPDB是很低的负值,应为硫酸盐还原、甲烷氧化或喜氧喜盐微生物白云岩。如果δ 13CPDB比-10‰还低,则为甲烷氧化微生物白云岩。甲烷生成微生物白云岩的δ 13CPDB通常都是正值,与无机成因的泥粉晶白云岩的δ 13CPDB有重合,但如果δ 13CPDB在3‰以上,则通常是甲烷生成微生物白云岩。
-
硫酸盐还原微生物白云岩、甲烷氧化微生物白云岩和甲烷生成微生物白云岩都形成于缺氧还原环境,其颜色普遍为深灰、灰黑等还原色,而且普遍含准同生期的自生黄铁矿等还原性自生矿物。而喜氧喜盐微生物白云岩形成于富氧环境,其颜色总体较浅,可为红色、黄褐色等氧化色,可含氧化性自生矿物,如赤铁矿、褐铁矿等。因此,根据颜色和自生矿物有可能将喜氧喜盐微生物白云岩与硫酸盐还原微生物白云岩、甲烷氧化微生物白云岩和甲烷生成微生物白云岩区分开来,但不能区分硫酸盐还原微生物白云岩、甲烷氧化微生物白云岩和甲烷生成微生物白云岩。
-
硫酸盐还原微生物白云岩、甲烷生成微生物白云岩和喜氧喜盐微生物白云岩的形成都需要沉积物富含有机质(腐泥型)。甲烷氧化微生物白云岩虽然不直接利用有机质,但甲烷通常是由有机质分解产生的。因此,所有微生物白云岩都直接或间接与盛产有机质的沉积环境有关,与富含有机质的沉积物互层共生。
海(湖)环境中的有机质主要是由水生低等植物(藻类等)产生的。由于植物生长需要阳光,因此水体要浅,并且环境条件恶劣(如滞流缺氧),不利于以植物为食的水生动物生存。潟湖、海湾(湖湾)是有利于植物大量发育的环境。一些深海(深湖)区,如果浮游生物大量发育且死后沉落到水底,其沉积中可富含有机质。潮坪上如果发育藻席,其沉积中有机质含量可以较高。
硫酸盐还原微生物白云岩、甲烷氧化微生物白云岩和甲烷生成微生物白云岩都形成于缺氧还原环境,因此与缺氧还原环境沉积(如滞流环境、深水环境)互层共生。硫酸盐还原微生物白云岩的形成需要水中含有较多的硫酸盐,因此通常只出现在海相沉积中,一般不会出现在湖泊沉积中,因为湖泊中硫酸盐含量通常很低。甲烷氧化微生物白云岩和甲烷生成微生物白云岩则无此限制。喜氧喜盐微生物白云岩形成于氧化环境,因此与浅水或暴露的氧化环境沉积(如潮坪)互层共生。
-
有序度是白云石晶体结构研究中重要参数之一,理想的白云石晶体为三方晶系,晶体结构中的阳离子Ca2+和Mg2+是按1∶1完全有序排列;受到白云石化成因、温度等因素的影响,白云石有序度通常无法达到理想形态而呈现不同的有序度[48]。由于微生物的参与使Ca2+和Mg2+排列不规则,导致微生物白云岩矿物有序度值较低,为0.6左右[49]。同时,在微生物白云石中可以探测到晶胞有机质。
2.1 晶粒大小方面的标志
2.2 晶体形态方面的标志
2.3 碳氧同位素方面的标志
2.4 颜色和自生矿物方面的标志
2.5 共生组合方面的标志
2.6 晶体结构方面的标志
-
现在关于微生物形成白云岩的解释是:微生物通过提高
另外,国外学者大多是在实验室微生物培育和现代沉积研究的基础上提出来的微生物介导白云岩模式,由于地质历史中的微生物与现代种类相差巨大,微生物本身也发生了进化,要从古代巨厚的白云岩中找到微生物的成因,则需沉积学家更多更深入的研究。总之,关于微生物如何形成白云岩的谜仍未完全解开,尚需进一步更深入、广泛的研究。
-
上述微生物白云岩的形成机理都没有涉及这样一个问题:微生物白云岩是原生还是次生交代的?
从形成机理上看应为原生的,因为白云石是在微生物作用帮助下沉淀出来的。但现代实例中的资料显示,微生物白云岩应该是次生交代的,因为白云石的含量从表层向下逐渐升高,最终达100%,而钙质矿物则逐渐减少,直至为0。这说明微生物白云岩的形成伴随着钙质矿物的溶解。
上述无论哪种微生物白云岩的形成机理,其现代实例都是准同生期形成的白云岩。白云石是从沉积物孔隙水中形成的,而非直接从沉积水体沉淀的。根据样品的扫描电镜观察发现,白云石虽然是在沉积物孔隙中沉淀的,但未见交代高镁方解石(或文石)的标志。微生物白云岩很可能是通过溶解—沉淀过程形成的,即高镁方解石溶解,微生物白云石在附近沉淀,而不是在高镁方解石溶解后腾出的位置沉淀。这种溶解—沉淀在微观上不是严格的交代作用,因为白云石晶体并没有占据高镁方解石溶解后腾出的位置;但宏观上表现为交代作用,因为白云岩确实占据了石灰岩的位置。

 DownLoad:
DownLoad: