
HTML
-
沉积有机质和原油中的生物标志化合物或称“分子化石”,具有相当稳定的结构,在地质热演化历史中可以保留从生物先体中继承下来的基本碳骨架特征,最终形成稳定的地质结构[1]。生物标志化合物可以用于判断有机相,热成熟度以及沉积环境和生物来源[2-3]。4-甲基甾烷,甲藻甾烷和三芳甲藻甾烷是沟鞭藻在古代沉积物中重要的存在形式。C30-4-甲基甾烷是常见的4-甲基甾烷系列,前人研究认为其有两种来源,一种是来源于沟鞭藻中的4-甲基甾醇,另一种是来源于细菌(产甲烷菌,荚膜甲基球菌)合成的4-甲基甾醇[4-7]。C30-4-甲基甾烷首次发现于海相沟鞭藻的沉积物中,古代淡水湖相中沟鞭藻主要合成4α-甲基-24-乙基甾醇而非甲藻甾醇[3,7]。黄第藩等[4]在咸化湖泊中检测到不同构型的4-甲基甾烷,认为4-甲基甾烷不一定仅来源于沟鞭藻,也许微生物起着更为重要的作用。另外,前人在海相和湖相环境中均检测到甲藻甾烷,其中半咸水和咸水湖相环境中的含量极高[7-10]。三芳甲藻甾烷是甲藻甾烷逐步芳构化的结果,几乎专一性地来自沟鞭藻中的甲藻甾醇,可以作为指示沟鞭藻的分子化石。Moldowan et al.[11]认为前寒武纪至白垩纪三芳甲藻甾烷与沟鞭藻和疑源类化石具有明显的相关性;张水昌等[12]在寒武系和上奥陶统中发现了高丰度的三芳甲藻甾烷,认为其来源于沟鞭藻;王广利等[13-14]利用三芳甲藻甾烷指数(TDSI)来表征三芳甲藻甾烷与3-甲基三芳甾烷的相对含量,认为三芳甲藻甾烷的丰度与沟鞭藻的丰度密切相关,且与姥鲛烷/植烷(Pr/Ph)、伽马蜡烷指数等参数无明显相关性,受沉积环境的影响较小[13-15]。
北部湾盆地古近系始新统流二段富有机质页岩中检测到三芳甲藻甾烷和高丰度的C30-4-甲基甾烷,这些页岩中富含大量藻类化石,如沟鞭藻(甲藻类)、盘星藻、葡萄藻和光面球藻等[16],这对该段有机质的富集作出了重要贡献。本次研究通过对北部湾盆地烃源岩中甾类分子化石的对比研究,结合古沉积环境探讨甲藻甾烷,三芳甲藻甾和C30-4-甲基甾烷分布特征及其与沟鞭藻的成因关系。
-
北部湾盆地位于中国南海北部大陆边缘,是早古近纪随南海盆地向南扩张形成的陆相新生代裂谷盆地[17-18]。盆地经历了古近系裂陷期和新近系裂后坳陷期两个构造演化阶段,古新世早期开始沉降[19],古近系的陆相沉积,划分为长流组、流沙港组和涠洲组;新近系的海相沉积,划分为下洋组、角尾组、灯楼角组和望楼港组。北部湾盆地的烃源岩发育在古近系始新统流沙港组和渐新统涠洲组。其中流沙港组自下而上划分为流三段(El3)、流二段(El2)和流一段(El1),流二段沉积时期是构造作用最强烈的时期,主力烃源岩也在该段发育,流二段处于湖盆扩张期,由于沉降速率大,古湖泊水域扩大,形成欠补偿沉积。由于该时期水体存在较好的分层结构,水体中存在大量藻类等水生生物且古湖底层缺氧而导致沉积有机质得到有效保存[16],发育大套中—深湖相暗色泥岩、页岩。近年来勘探研究表明,流二段顶部富有机质页岩为7~25 m,底部富有机质页岩厚度总体大于40 m,分布范围广,厚度较稳定,是该盆地中的优质生油岩[16,20-21]。
本文研究的样品取自北部湾盆地涠西南凹陷和乌石凹陷古近系流沙港组二段,共19件源岩样品,分别为油页岩、泥岩和页岩。油页岩生烃潜力明显优于中深湖相泥岩和页岩,大部分处于低熟—成熟阶段,油页岩有机质类型主要为Ⅰ~Ⅱ1型,腐泥组组分含量最高,矿物沥青基质含量高,富含次生显微组分,油页岩中腐泥组+壳质组组分含量略高于泥岩和页岩。深湖相泥岩和页岩有机质类型为Ⅱ1~Ⅱ2型。不同类型烃源岩倾油气性存在一定差别,但均以倾油性为主。
-
烃源岩样品粉碎至80目后,用二氯甲烷进行索氏抽提24 h,抽提物脱沥青质后,在硅胶—氧化铝层析柱上用石油醚淋洗出饱和烃组分,用二氯甲烷:石油醚=2∶1(v∶v)淋洗出芳烃组分。在安捷伦GC7890A-5977型气相色谱/质谱联用仪上,使用HP-5MS型毛细色谱柱(60 m×0.25 mm×0.25 μm),进行色谱—质谱(GC-MS)分析。饱和烃升温程序的初始温度为50 ℃,保持1 min,以20 ℃/min的速率,升至120 ℃,以4 ℃/min的速率升至250 ℃,再以3 ℃/min的速率升至310 ℃,恒温30 min;芳烃升温程序的初始温度为80 ℃,保持1 min,以3 ℃/min的速率升至310 ℃,恒温20 min。以氦作载气,分流比20∶1,流量1 mL/min。离子化方式为电子轰击(EI),离子源温度230 ℃,四极杆温度150 ℃,电子轰击能量70 eV,获取数据扫描范围50~580 amu,扫描方式为全扫描(Scan)或选择离子(SIM)检测。
1.1. 地质背景及样品
1.2. 实验方法
-
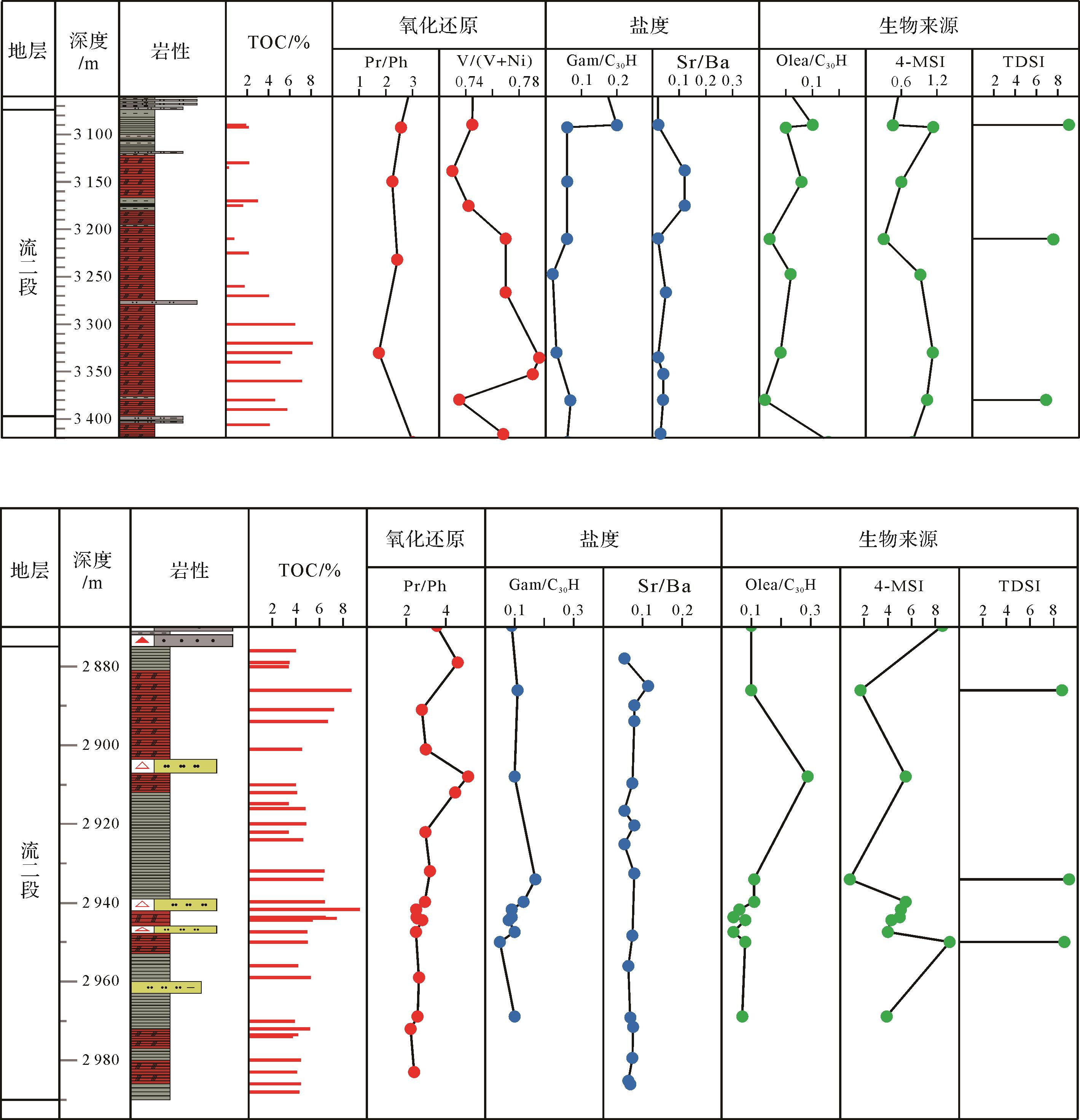
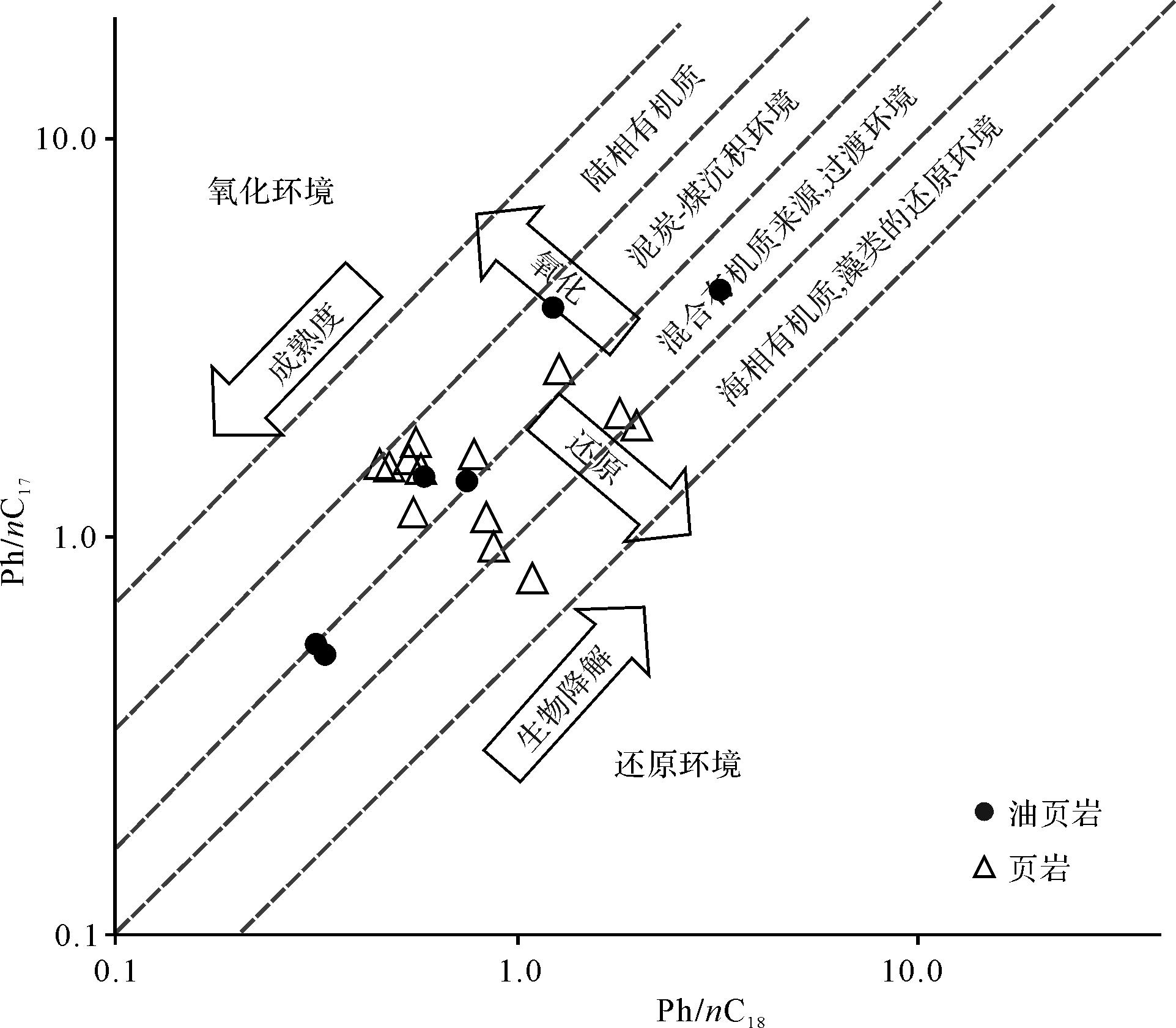
沉积环境受水深、温度、盐度、氧化还原性以及水动力条件的影响而不同,流沙港组二段主要为中—深湖相沉积,Huang et al.[22]认为流二段沉积时期水体缺氧、分层,且具有一定的矿化度。姥鲛烷和植烷可以判断原始沉积环境的氧化还原性,二者在现代沉积物、古代沉积岩和原油中广泛分布,来源于藻类、高等植物中的叶绿素和藻菌类中植基侧链形成的植醇,在不同的氧化还原条件下向不同方向转化,在弱氧化酸性条件下易形成姥鲛烷,而在还原碱性条件下经脱水作用易形成植烷。流二段烃源岩姥鲛烷和植烷比值(Pr/Ph)分布范围变化不大,Pr/Ph分布范围为0.85~4.23(表1),平均2.17,总体介于1~3,个别大于3;Pr/nC17值分布在0.51~4.15的范围内,平均值为1.67,Ph/nC18值分布在0.32~3.2的范围内,平均值为0.96(图1),表明形成于偏还原的沉积环境。V/(V+Ni)也可以反映氧化还原条件,V/(V+Ni)>0.77时为缺氧、极贫氧环境,V/(V+Ni)介于0.60~0.77时为贫氧、次富氧环境,V/(V+Ni)<0.60时为富氧环境[23]。本次所研究的样品,流二段烃源岩V/(V+Ni)值分布在0.73~0.80的范围内,表明其形成于次富氧的偏还原沉积环境(图2)。
编号 井名 深度/m 岩性 TOC/% Pr/Ph Gam/C30H 4MSI TDSI 1 WA11-2-1 3 350 油页岩 6.34 1.49 0.12 0.78 0.23 2 WA11-2-2 3 018 油页岩 4.26 1.48 0.11 1.33 0.16 3 WA12-11-1D 2 326 油页岩 5.31 3.21 0.03 0.57 0.50 4 WA12-11-1D 2 644 油页岩 6.47 2.67 0.03 0.42 0.33 5 WA6-3S-1 3 380 油页岩 4.58 1.59 0.07 1.03 0.24 6 WA6-3S-1 3 490 油页岩 7.52 1.49 0.15 1.33 0.15 7 WB16-1-2 2 598 页岩 0.97 4.23 0.07 0.16 0.62 8 WB1-6-1 1 480 页岩 3.34 1.54 0.27 0.20 0.47 9 WA11-2-2 2 720 页岩 2.08 3.15 0.05 0.25 0.39 10 WB16-1-5 2 202 页岩 1.63 1.44 0.09 0.07 0.50 11 WB16-1-5 2 632 页岩 1.40 2.47 0.10 0.13 0.54 12 WB16-1-3 2 351 页岩 1.87 0.85 0.07 0.12 0.43 13 WB16-1-3 2 609 页岩 1.39 1.20 0.08 0.13 0.45 14 WB16-6-1 1 932 泥岩 1.74 2.22 0.05 0.69 0.43 15 WB16-8-1 2 886 页岩 4.34 2.69 0.06 0.69 0.46 16 WB16-8-1 2 934 页岩 3.15 3.31 0.06 0.49 0.52 17 WB16-8-1 2 950 页岩 2.51 2.17 0.07 0.28 0.50 18 WA6-3S-1 3 090 页岩 1.92 1.53 0.20 0.45 0.51 19 WA6-3S-1 3 210 页岩 0.80 2.89 0.06 0.31 0.37 
Figure 1. Correlation diagram of Pr/nC17 and Pr/nC18 in Second member of Liushagang Formation, Beibuwan Basin

Figure 2. Geochemical profile of source rock in Second member of Liushagang Formation, Beibuwan Basin
伽马蜡烷是一种五环三环萜烷中的非藿烷化合物,最早发现于绿河页岩中,其前身物被认为是四膜虫醇,存在于某些原生动物、光合细菌和一些可能的其他生物的细胞膜中[11,24-26]。Kleemann et al.[27]研究认为伽马蜡烷不仅可以指示沉积水体的盐度,也可以指示水体的分层事件。沉积环境为淡水时,伽马蜡烷含量低,随水体盐度增加,伽马蜡烷含量随之增加。流二段烃源岩的伽马蜡烷含量均较低,伽马蜡烷指数(Gam/C30H)值绝大多数小于0.2,表明水体盐度不高,总体为淡水—微咸水沉积。Sr/Ba值可判断海相或陆相沉积,与Sr相比,Ba的化合物的溶解度要低,因而多数Ba在近岸沉积物中富集,仅有少量钡进入海中,且Sr的迁移能力要高于Ba,可能迁移到大洋深处[28]。前人研究认为,海相沉积Sr/Ba>1,陆相沉积Sr/Ba<1[29-30]。Sr/Ba值也可以判断古湖泊水体盐度,Sr/Ba<0.5为淡水环境,Sr/Ba介于0.5~1.0为半咸水环境,Sr/Ba>1.0为咸水环境[30-31]。研究区流二段烃源岩所分析的Sr/Ba大都小于1,其值分布在0.02~0.12,总体为淡水湖相沉积(图2)。综合来看,流二段为淡水—微咸水沉积环境。
-
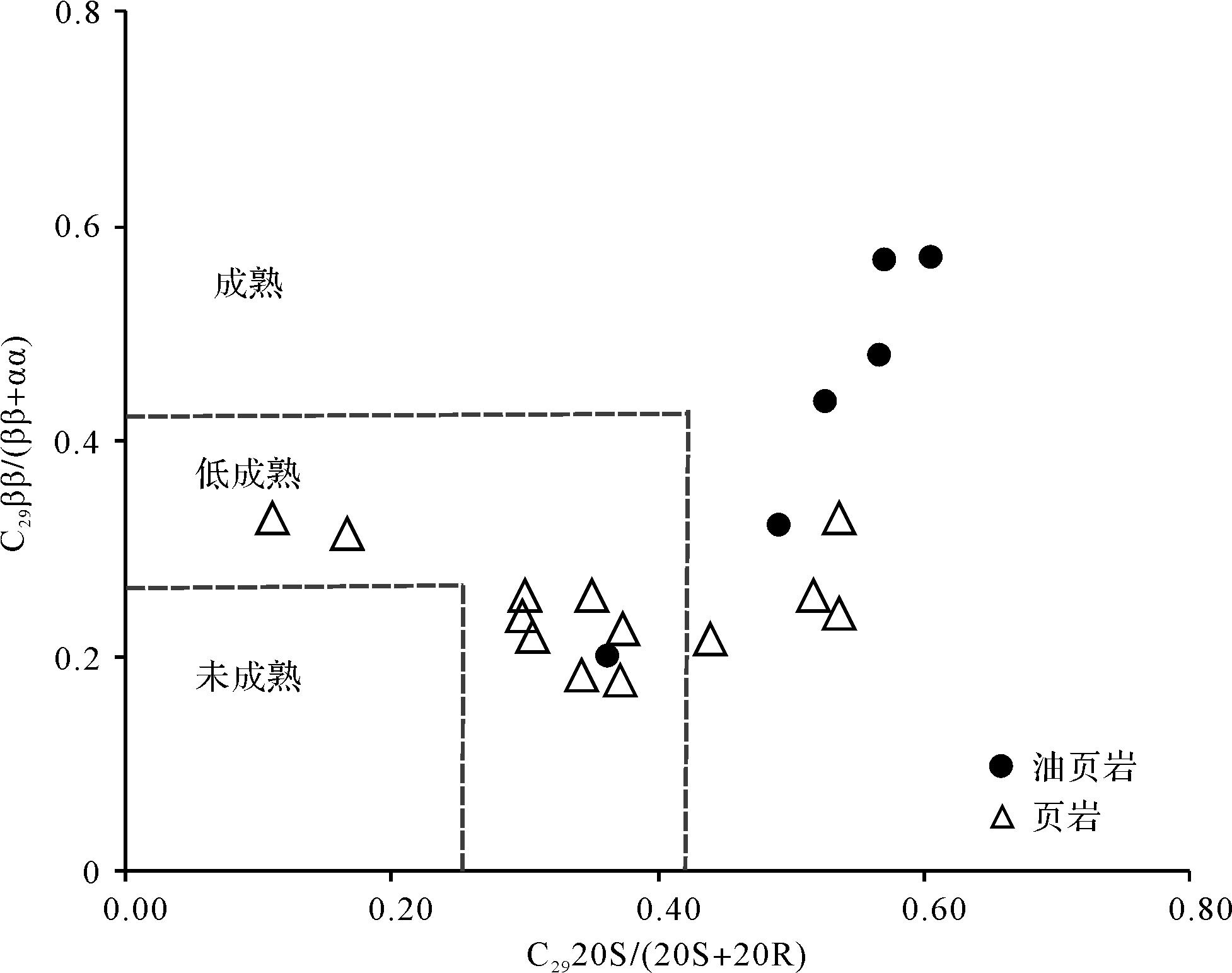
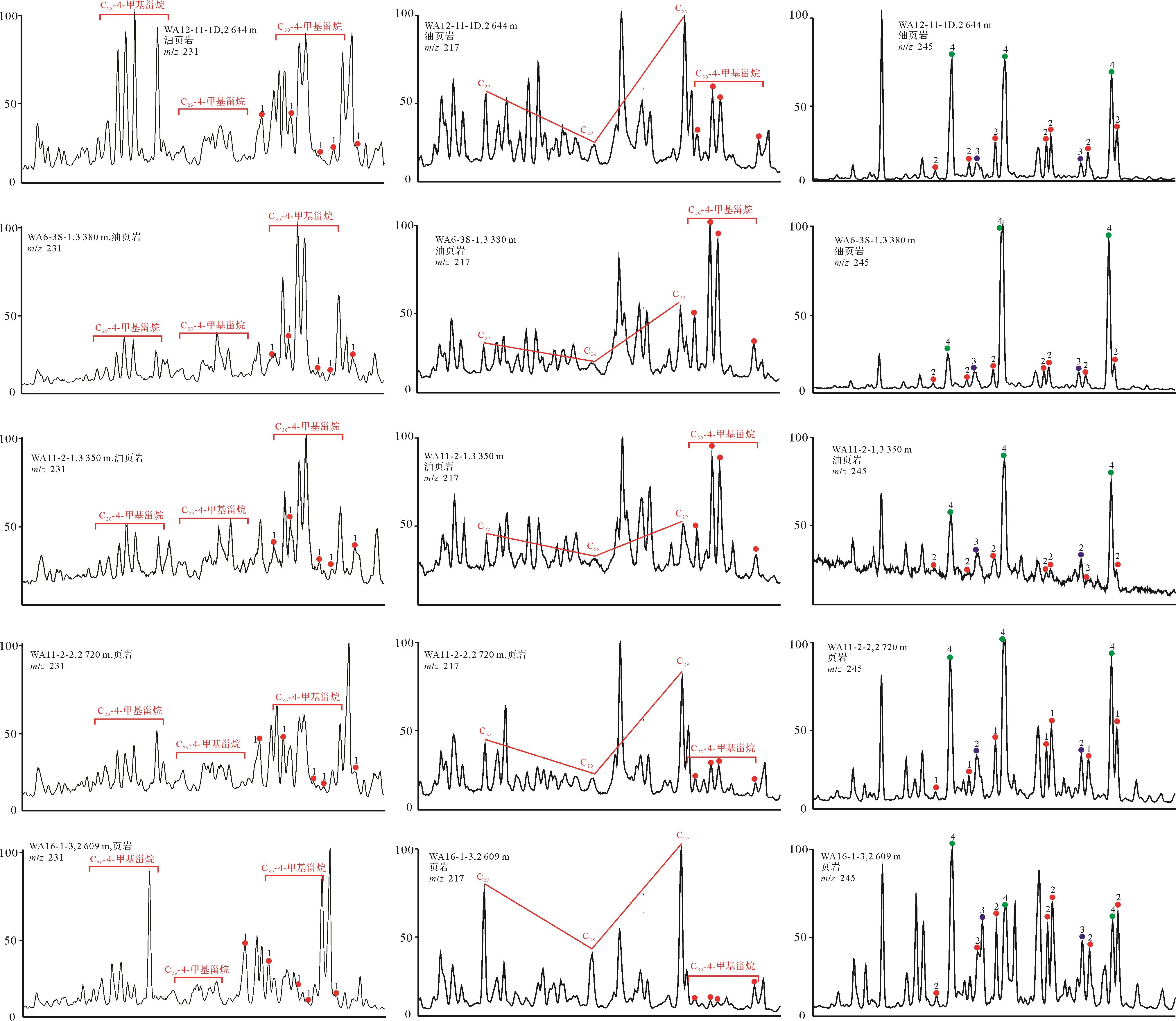
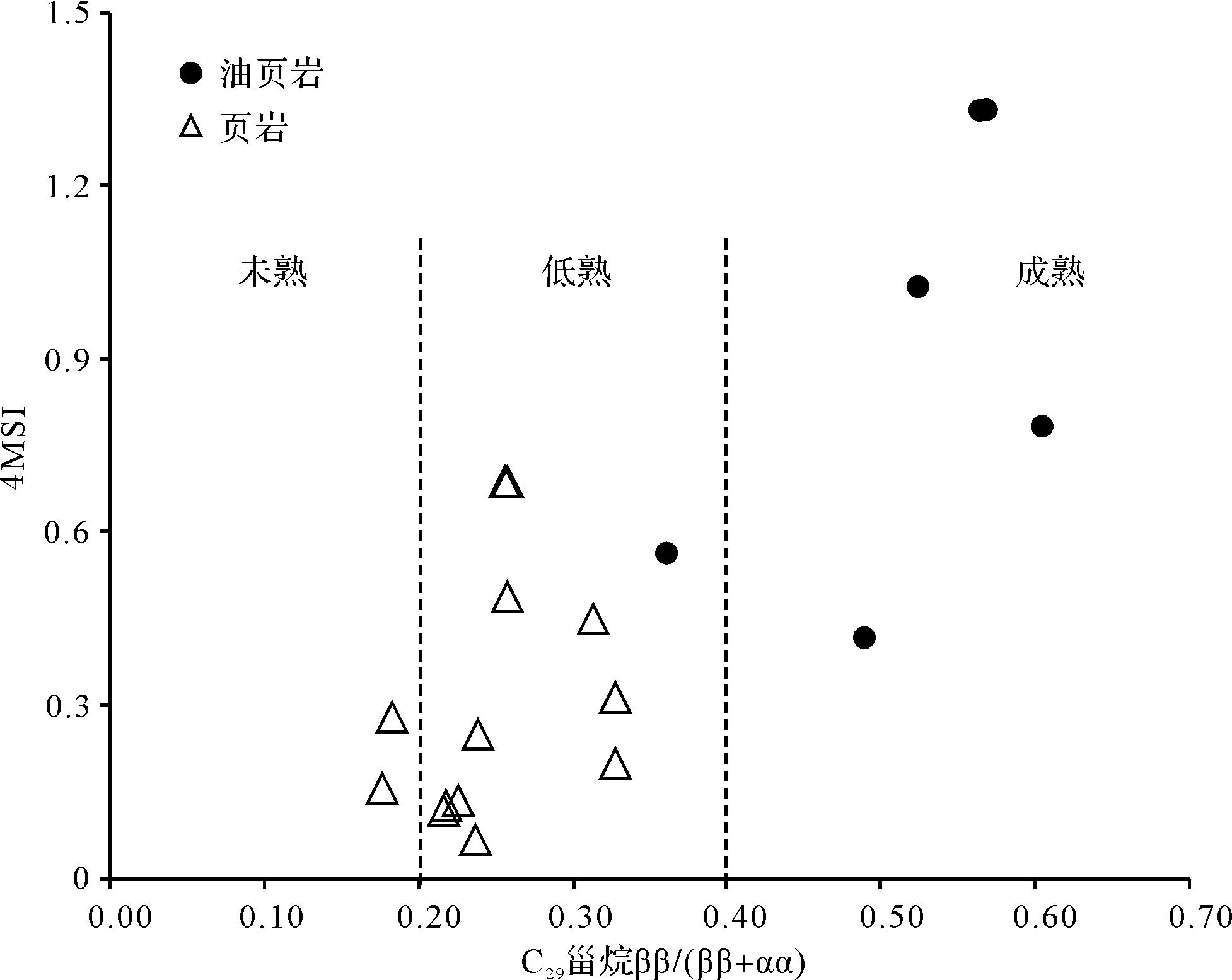
北部湾盆地流二段烃源岩中,C27、C28与C29规则甾烷多为不对称“V”型,C27/C29规则甾烷介于0.40~1.21,平均值为0.72,表明该时期既有低等水生生物来源也有陆源高等植物来源,以水生生物尤其是藻类为主。盆地烃源岩总体处于低成熟—成熟阶段(图3)。另外,研究区也检测到C28—C30-4-甲基甾烷系列,但C28-和C29-4-甲基甾烷的相对丰度远低于C30-4-甲基甾烷。中—深湖相沉积层中以高丰度的C30-4-甲基甾烷为主要特征,流二段油页岩中最为丰富(图4),4MSI(C30-4-甲基甾烷/C29规则甾烷)的平均值达到0.9以上(表1),C30-4-甲基甾烷除了4α-和4β-两种构型外,还有其他一些不明确的结构。4-甲基甾烷有两种来源,一种可由甲藻(沟鞭藻)提供,另一种则类源于细菌[4-5,32]。

Figure 3. Correlation diagram of C29ββ/(ββ+αα) and C2920S/(20S+20R) in Second member of Liushagang Formation, Beibuwan Basin

Figure 4. Correlation diagram of triaromatic dinosteroid hydrocarbon index (TDSI) and 4⁃methyl sterane index (4MSI) in the Second member of the Liushagang Formation, Beibuwan Basin
同时,研究区油页岩、泥岩、页岩中检测到甲藻甾烷和三芳甲藻甾烷,但二者的相对丰度均相对较低(图4、表1),由于三芳甲藻甾烷与3-甲基三芳甾烷的丰度相当,难以通过计算显示其丰度差异,因而在前人使用的TDSI指数[14]的分母中加上了与三芳甲藻甾烷丰度差别较大的4-甲基三芳甾烷。TDSI=三芳甲藻甾烷/(三芳甲藻甾烷+3-甲基三芳甾烷+4-甲基三芳甾烷),该值介于0.15~0.62,平均值为0.41(图4)。三芳甲藻甾烷由甲藻甾烷逐步芳构化而来[12],几乎专一性地由沟鞭藻体内的甲藻甾醇提供。
-
前人所检测到的C30-4-甲基甾烷主要出现在海相环境或者咸化湖泊中,在柴达木盆地第三系盐湖及渤海湾盆地始新世湖相中均检测到一定数量的C30-4-甲基甾烷[33],前人认为海相或咸化湖泊中存在能够提供4-甲基甾烷先质的沟鞭藻。而北部湾盆地处于弱还原的淡水—微咸水湖泊环境,在油页岩中检测到高丰度的C30-4-甲基甾烷,页岩中检测到中—低丰度的C30-4-甲基甾烷,二者中同时检测到较低丰度的甲藻甾烷和三芳甲藻甾烷,三者都被认为可以由沟鞭藻提供,且甲藻甾烷和三芳甲藻甾烷几乎专一性地由沟鞭藻提供。至于细菌是否也参与了C30-4-甲基甾烷的形成,国内外许多学者也开展了一定的探究。Bird et al.[34]曾检测到一种甲烷细菌含有4,4-二甲基、4-甲基与4-脱甲基甾醇,虽然这些细菌产生4-甲基甾醇,但尚未明确是否能合成C30化合物的23,24-二甲基或24-乙基取代基,只合成了C28-4α甲基甾醇和C29-4,4-二甲基甾醇[9]。黄第藩等[4]认为柴达木第三系盐湖相沉积中的4-甲基甾烷主要是成岩作用中甲烷生成菌活动的产物,但其环境条件必须缺氧、还原、富硫酸盐,并且地温低于85 ℃,酸碱性为中性(pH=6.4~7.5)。很显然,结合本区弱还原且盐度低的环境条件以及低丰度的C28-和C29-4-甲基甾烷分布,C30-4-甲基甾烷几乎不可能是由细菌提供的。
沟鞭藻的种类繁多,在海洋和淡水环境中均有分布,但其组合的面貌有差异,非海相的半咸水和过渡相的沟鞭藻组合体现了典型海、陆相之间的过渡特征。海相环境中检测到许多沟鞭藻的种属,如Peridinium foliaceum。陆相淡水环境中也有许多典型的沟鞭藻种属,如盘星藻属(Pediastrum),拉氏多甲藻属(P.lomnickiiWoloszynska),该种属主要提供4α-甲基甾醇[35-37]。济阳凹陷咸化湖泊中以德弗兰藻属(Deflandrea)为主,另外还有多刺甲藻属—菱球藻属(entusidinium-Pararhombodella),渤海藻类等。前人研究发现,不同种属的沟鞭藻都可以提供4-甲基甾醇和甲藻甾醇[5-7],北部湾盆地早第三纪湖泊沉积中既有盘星藻、葡萄藻等淡水浮游藻类,也有属于非海相沟鞭藻的百色藻和属于疑源类的球藻以及绿藻[8]。因而,沟鞭藻种属的不同不是造成C30-4-甲基甾烷、甲藻甾烷和三芳甲藻甾烷差异性分布的原因。这使得我们需要考虑沉积环境以及有机质沉降过程中成岩作用的影响。
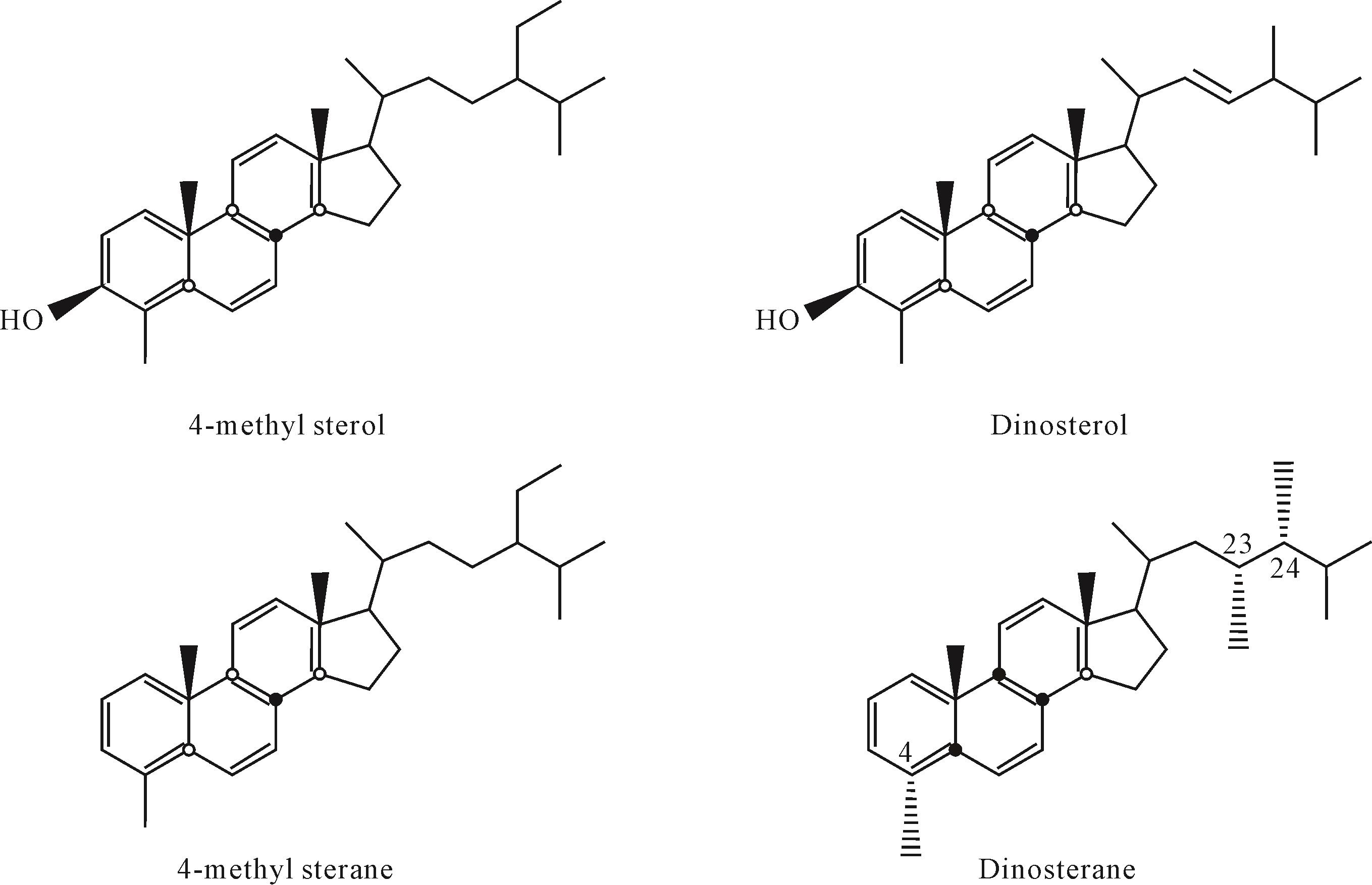
4-甲基甾烷和甲藻甾烷以及它们的先质在结构上均不同(图5),北部湾盆地绝大多数烃源岩和原油中所检出的C30-4-甲基甾烷均为24-乙基构型,即其侧链的C-24位上被一个乙基所取代,这种构型的C30-4-甲基甾烷在我国湖相沉积物及其相关原油中广泛发育,而甲藻甾烷在其侧链C-23和C-24位上的一个氢分别被一个甲基取代。Summons et al.[9]对三叠系—第三系原油和沉积物进行了研究,发现具有海相成因的原油和沉积物包含甲藻甾烷和4-甲基,24-乙基胆甾烷,而在白垩系—第三系湖相原油和沉积物中,甲藻甾烷缺失或丰度极低,4-甲基,24-乙基胆甾烷丰度高。渤海湾盆地沙河街组陆相咸化水体中高丰度的甲藻甾烷除了与沟鞭藻有关之外,还可能与海侵作用有关[38-39]。松辽盆地发生过海侵的青山口组一段和嫩江组一段、二段有含量相对较高的甲藻甾烷,且盐度越高,海侵的程度越大时甲藻甾烷含量越高,与海相具有相似生态环境的陆相半咸水—咸水环境中也存在大量沟鞭藻,能够形成丰富的甲藻甾烷[40],并且能够很好地保存下来。而北部湾盆地是一个与上述环境截然不同的典型陆相淡水—微咸水湖盆,水体盐度低且未遭受海侵,整体为弱还原的条件,因而没有大量甲藻甾烷形成和保存下来。同样来源于沟鞭藻的C30-4-甲基甾烷在该盆地却有着很高的丰度,可能是由于其先质不同,4-甲基甾醇比甲藻甾醇少一个不饱和双键(图6),反应相对较易进行,因而在成岩过程中甲藻甾醇需要更加还原的环境来加氢还原形成甲藻甾烷,而4-甲基甾醇在相对弱还原的环境中就可以形成C30-4-甲基甾烷。

Figure 5. Mass chromatograms of steroids in the Second member of Liushagang Formation, Beibuwan Basin

Figure 6. Structural formulae of 4⁃methyl sterane, dinosterane and their precursors
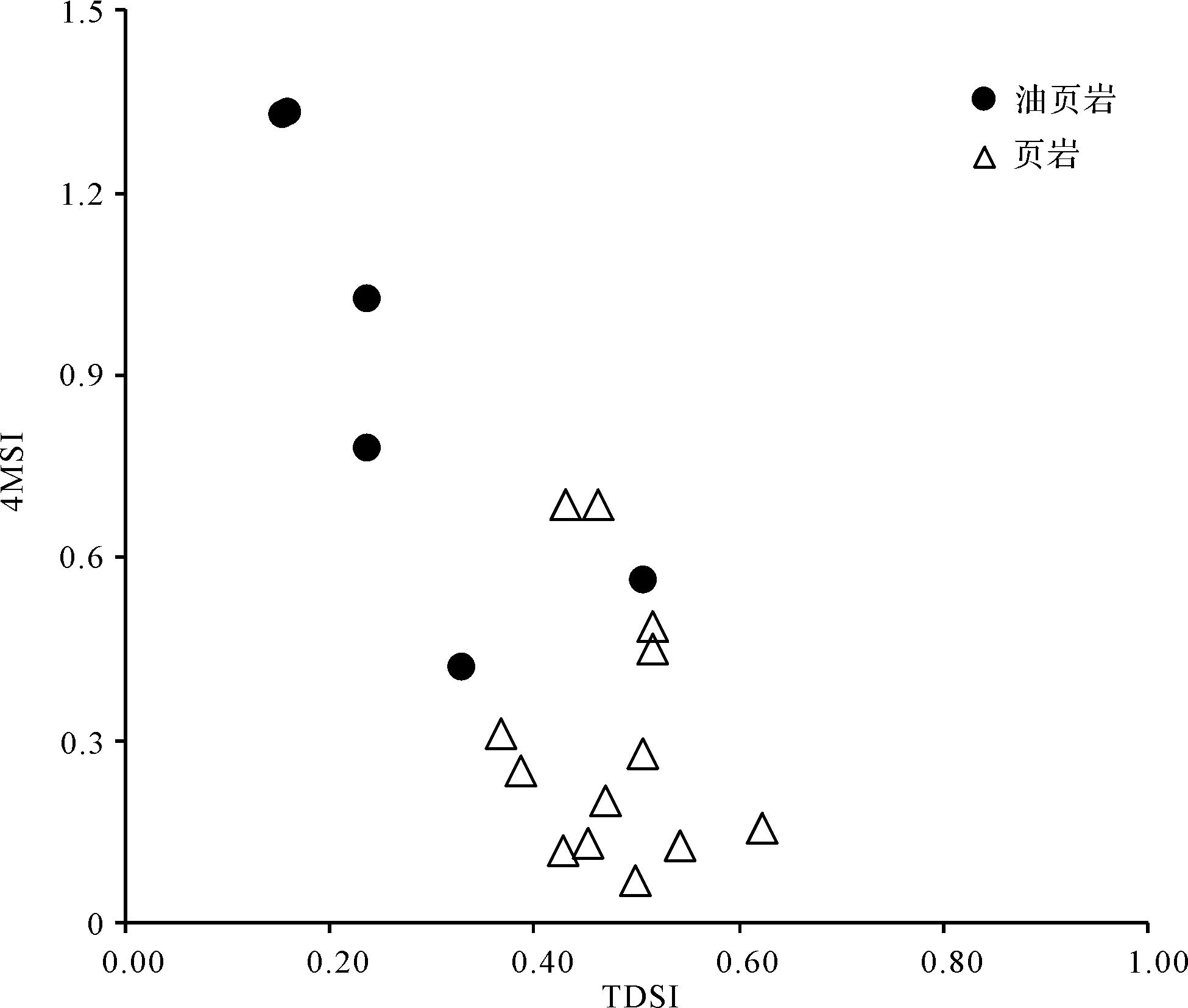
另外,结合其他成熟度参数研究了C30-4-甲基甾烷在有机质成烃过程中的变化(图7)。在有机质成岩作用和深成作用阶段,C30-4-甲基甾烷相对丰度的变化规律较为明显,随着成岩作用加深,C30-4-甲基甾烷相对丰度明显增加,这一现象也说明沟鞭藻类的输入量不完全是决定C30-4-甲基甾烷相对丰度的因素,而与成岩作用过程有着较为密切的关系。盆地油页岩中高丰度的C30-4-甲基甾烷与其相对较高成熟度也有一定关系。

Figure 7. Correlation diagram of 4⁃methyl sterane index (4MSI) and maturityin the Second member of the Liushagang Formation, Beibuwan Basin
2.1. 流沙港组二段沉积环境分析
2.2. 甾类分布特征与生源构成
2.3. 成因分析
-
(1) 北部湾盆地流二段烃源岩中藻类丰富,油页岩中检测到高丰度的C30-4-甲基甾烷,页岩中检测到中—低丰度的C30-4-甲基甾烷,油页岩和页岩中同时检测到低丰度的甲藻甾烷和三芳甲藻甾烷。
(2) 北部湾盆地流二段烃源岩形成于弱还原的淡水—微咸水湖泊环境。这种沉积条件不适宜甲藻甾烷和三芳甲藻甾烷的形成,而可能形成高丰度的C30-4-甲基甾烷。
(3) 北部湾盆地高丰度的C30-4-甲基甾烷主要来源于非海相的沟鞭藻。油页岩中C30-4-甲基甾烷相对丰度明显高于页岩,这与油页岩相对较高的成熟度有着密切的关系。另外,沟鞭藻种属的不同不是造成C30-4-甲基甾烷,甲藻甾烷和三芳甲藻甾烷差异性分布的原因,可能是由于其藻类产出先质不同,在弱还原的条件下,更易生成C30-4-甲基甾烷,而难以形成甲藻甾烷和三芳甲藻甾烷。

 DownLoad:
DownLoad: