
HTML
-
前人关于潮坪沉积环境的研究大多侧重于潮坪的物理化学沉积特征,包括粒度分析[1⁃2]、层理特征描述[3⁃6]、沉积年代分析[7]等,然而生物对环境的反应是极其敏感的,其与沉积作用之间相互作用相互影响。遗迹学正是基于发现和分析生物成因的沉积构造,研究现代和古代的生物遗迹,主要强调生物与环境相互影响,密不可分,为鉴定和解释沉积环境起到了不可替代的作用[8⁃13],同时也为油气储层的预测研究提供依据[14⁃15]。
21世纪以来,国外学者对现代生物遗迹的沉积环境分析、形态学描述以及与遗迹化石的对比研究更加深入[16⁃20]。而潮坪现代生物遗迹的研究除了系统描述沉积学特征和遗迹学外[21⁃22],也有学者深入研究了不同研究区潮间带生物遗迹群落的组成分布特征,并进行物理化学因素对其影响的分析[23⁃24]。研究发现环境因素,如能量条件、底层性质、粒度、沉积速率、盐度、浑浊度、有机碳含量及其他物理化学特性会影响沉积介质中生物遗迹的种类、大小和分布[25⁃27],而生物遗迹特征如遗迹的分布、分异度、形态变化以及遗迹的行为生态学等在一定程度上可以灵敏地反映沉积过程中的物理化学环境因素[18,24,28]。
黄河三角洲是一类典型的高泥沙高流量沉积环境,其沉积作用是物理、化学和生物三者相互影响的复杂动态过程,物理因素和沉积构造是了解现代三角洲沉积环境的有力工具[7,29⁃31],然而伴随沉积产生的生物遗迹的研究一般只是遗迹形态[32]、生物沉积构造、组成及分布特征[33⁃35]。现代潮坪遗迹学特征对物化因素的响应关系既可以指示沉积环境的变化,也可以为潮坪古遗迹学提供实证材料。因此,详尽且精细的黄河三角洲潮坪环境的生物遗迹特征与沉积过程中物理化学环境因素的响应,还有待系统研究。
-
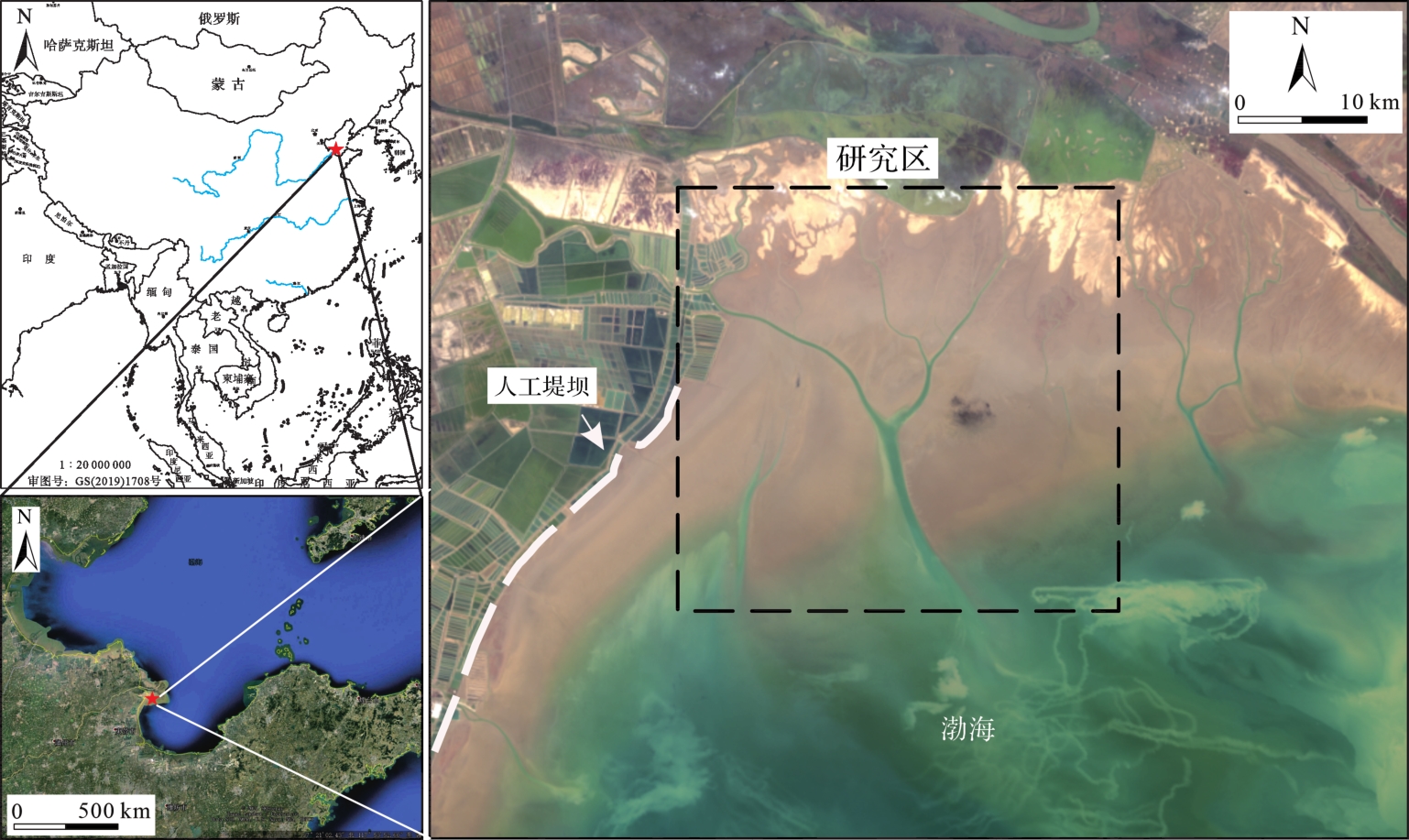
渤海湾黄河三角洲潮坪面积广阔,底栖生物丰富,主要受不规则的半日潮控制,平均潮差为0.73~1.77 m,平均速度为0.5~1.0 m/s[36]。潮坪表面有大量树枝状潮沟,微地形变化复杂。潮流为往复流,一般平行于海岸线,涨潮时研究区被海水淹没,落潮时露出水面,为野外工作提供了有利的条件。
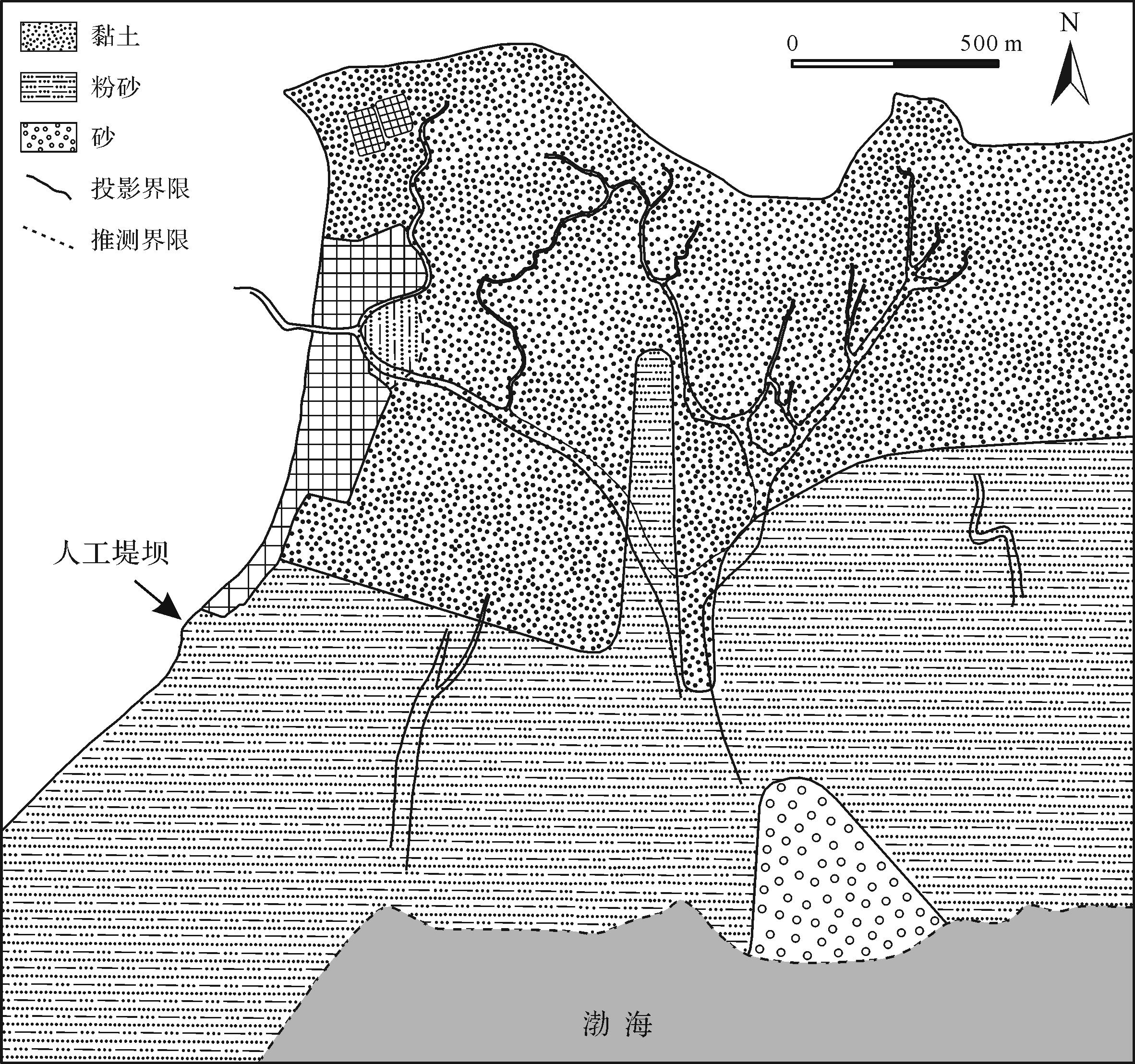
研究区位于山东省东营市黄河三角洲南部潮坪(119°0'~119°10' E,37°37'~37°43' N)(图1),坡度自岸向内陆缓慢增加,且涨潮流速大于落潮流速,因此由涨潮带来的沉积物很难再次搬运回去。潮坪环境依次为潮上带,潮间带以及潮下带[37],沉积物主要来源是随潮汐往复的黄河入海的悬浮泥沙[38]。

Figure 1. Location of our study area
-
本研究选取黄河三角洲南部潮坪的现代生物遗迹作为野外考察对象,包括形态学、沉积学和遗迹学的调查。通过手持全球定位系统(GPS)绘制地面地图,确定了32个大多平行或垂直于海岸线的采样点(图2),在沉积物或生物分布不均匀的地区额外采样。在各个采样点,测算每平方米生物密度,采集离层面20 cm深的沉积物样品进行粒度和总有机碳(TOC)含量分析。在大多数采样点采集水样,测定盐度和浑浊度。

Figure 2. Sample and station locations within the tidal flat of the Yellow River Delta
取约0.2 g风干的沉积物样品进行预处理,利用Mastersize2000型激光粒度分析仪测定粒度分布,测量误差小于1%。所得到的平均粒径值是利用毫米值的矩量法求得的算术平均值,为了便于比较,转换为phi值。
对采集的沉积物样品进行TOC测试。首先在105 ℃下进行24 h的初始干燥,以去除间隙水。取研磨土样2 g及30 mL的盐酸放入封闭的样品管,按照液体高度在试管壁上做标记,放入离心机离心5 min。静置后倒掉上部液体,按标记高度加入纯水,再次放入离心机去酸化4~5遍,用pH试纸测试,直至pH值为7。将去酸化后的土样置于烧杯中,放入60 ℃的烘箱,烘3天,每天10 h。每个样品各称取2个5 mg烘干土,土壤标准物质称取3 mg,用锡杯包裹,每隔10个测试样品放置2个标样,使用仪器Apollo9000,通过燃烧法测定沉积物中有机碳的含量。在燃烧损失分析期间,用精确到0.001 g的电子秤测定有机物燃烧造成的重量损失。
通过计算洞穴开口、箱式取样法及X射线计算机断层扫描技术(CT)综合确定生物扰动程度[39]。用PVC管选取了33个表面形态完整、具有代表性的生物潜穴进行扫描和三维重构,获取其内部结构的完整信息。CT扫描实验室为中石化胜利油田勘探开发研究院地化实验室,样品直径为7.5 cm,长度为10~15 cm,所用仪器为Sanying牌扫描仪,电压150 keV,电流30 mA。将扫描得到的图像用中国科学院南京地质古生物研究所X射线断层扫描实验室的软件VG studio Max进行三维重构,得到生物潜穴的立体图像。
-
黄河三角洲潮坪潮上带处于低能环境,发育沙泥互层的水平层理,是由涨落潮带来的沙质悬浮物与憩流的泥质沉积交互形成。树枝状的潮道两侧水流增强和减弱都有规律,沉积物来源丰富,伴生的物理沉积构造十分发育,有平行层理、软沉积变形(图3a)、潮汐韵律层理(图3b)、板状交错层理、丘状交错层理、波状层理和生物扰动变形层理。软沉积变形于涨潮期间在上部潮道的分岔口处形成[28,40⁃41],这部分沉积物以被水充分饱和的黏土和细粉沙为主,沉积物内部黏结力极低,因此涨潮时的潮道两侧容易形成沉积物变形。研究区的潮汐韵律层理是沙质与泥质沉积物的韵律互层,由潮流的周期性变化形成。生物扰动变形层理的规模取决于生物个体的大小,形态由生物体的行为特性决定。层面沉积物间歇性暴露干涸,发育泥裂。潮上带沉积物在较短时间内沉积,层面上很少保留潮汐痕迹。

Figure 3. Common sedimentary structures in the tidal flat of the Yellow River Delta
潮间带宽阔,物理沉积构造相对简单,发育沙泥互层的平行层理、丘状层理(图3c)、波状层理、生物扰动变形层理和软沉积变形。潮水流动方向大多垂直、平行或斜交岸线,因此该部分沉积物表面波痕丰富,常见浪成波痕(图3e)。远离潮道的潮间带水动力较弱,在稳定的环境下发育轻微泥裂(图3f)。
潮下带处于高能环境,潮流流量和速度大,使得沉积物快速沉积,且沉积物富含水分,产生丰富的软沉积变形(图3d)及生物扰动变形构造。受海洋重力流的影响,其软沉积变形中的沙泥比例较潮上带大,且分布不均匀。层面上有丰富的生物扰动痕迹。
-
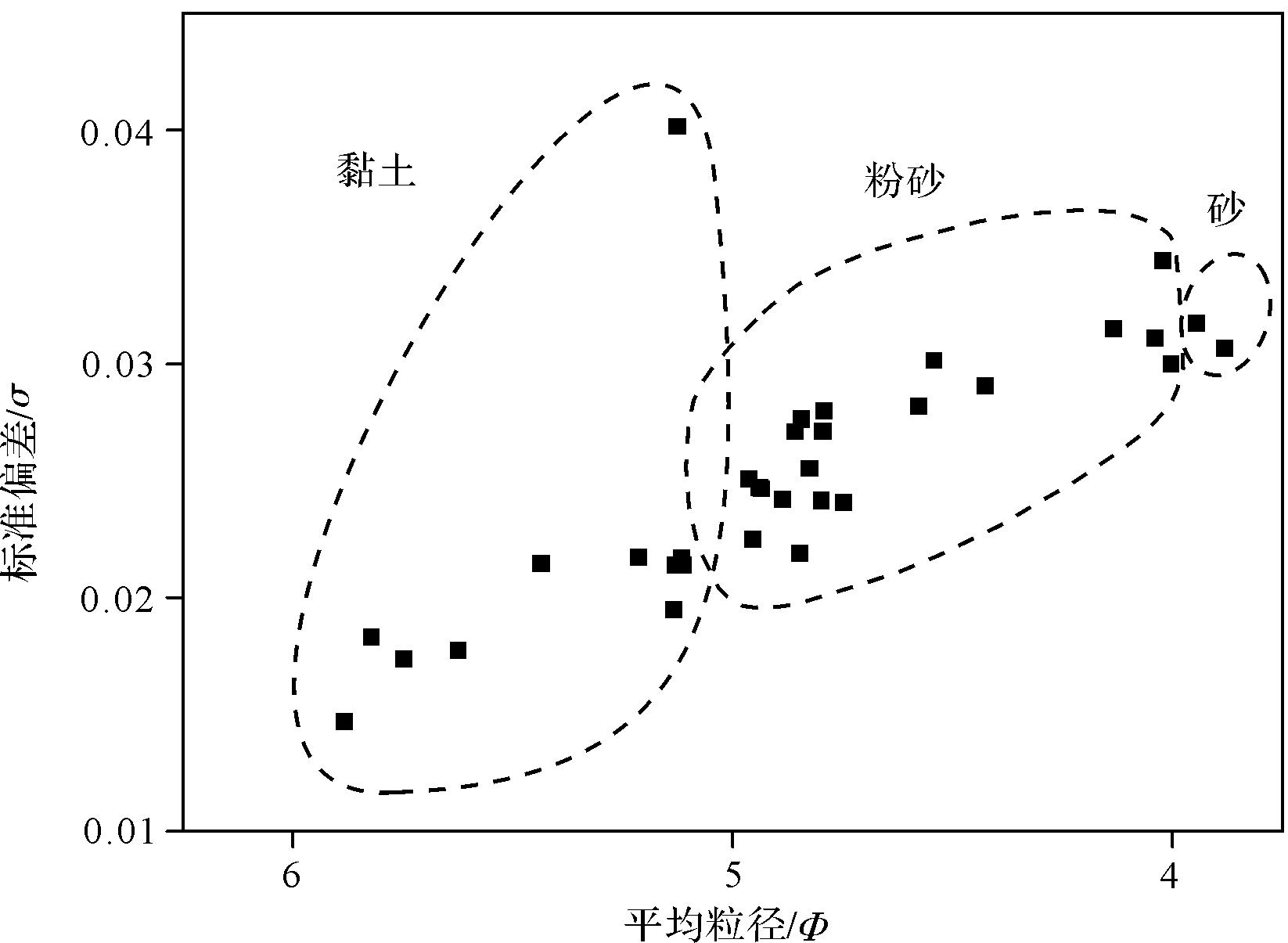
沉积速率是一种局部变化的应力,它与河流输入和泥沙粒径大小密切相关,沉积物的粒度越粗,基底的流动性就越低[42]。河流流量对沉积作用有强烈的影响,流量越大,可输送到海岸带的碎屑越大[23]。黄河三角洲潮坪沉积物多为细粒,以淤泥黏土和砂质粉沙为主,含少量的极细沙。粒级分布范围集中,主要粒径明显,分选系数介于0.01~0.04。按照传统分类方式[43⁃44],当分选系数小于0.35时,判定为极好(图4)。从潮下带到潮上带,沉积物粒度逐渐变小(图5),主要受潮汐和风的影响,颗粒大的砂质物质破碎,粗细混杂的物质经过较长距离搬运逐渐分异,砂质粉土和淤泥黏土在稳定的水动力条件下缓慢沉降形成。低潮线附近受急剧多变的海洋控制,波浪的活动及作用时间均比潮坪的较高部位强,因此粗粒沉积物随着潮汐的往复而逐渐沉降,以极细砂为主。研究区最北端长期处于暴露状态,水动力条件最弱,由于潮汐海水侵蚀以及强烈的蒸发形成盐碱地,可见石盐晶体、动物足迹以及稀疏的植被。

Figure 4. Mean grain size and sorting values for all tidal flat sediment samples collected at the Yellow River Delta

Figure 5. Sediment distribution within the tidal flat of the Yellow River Delta
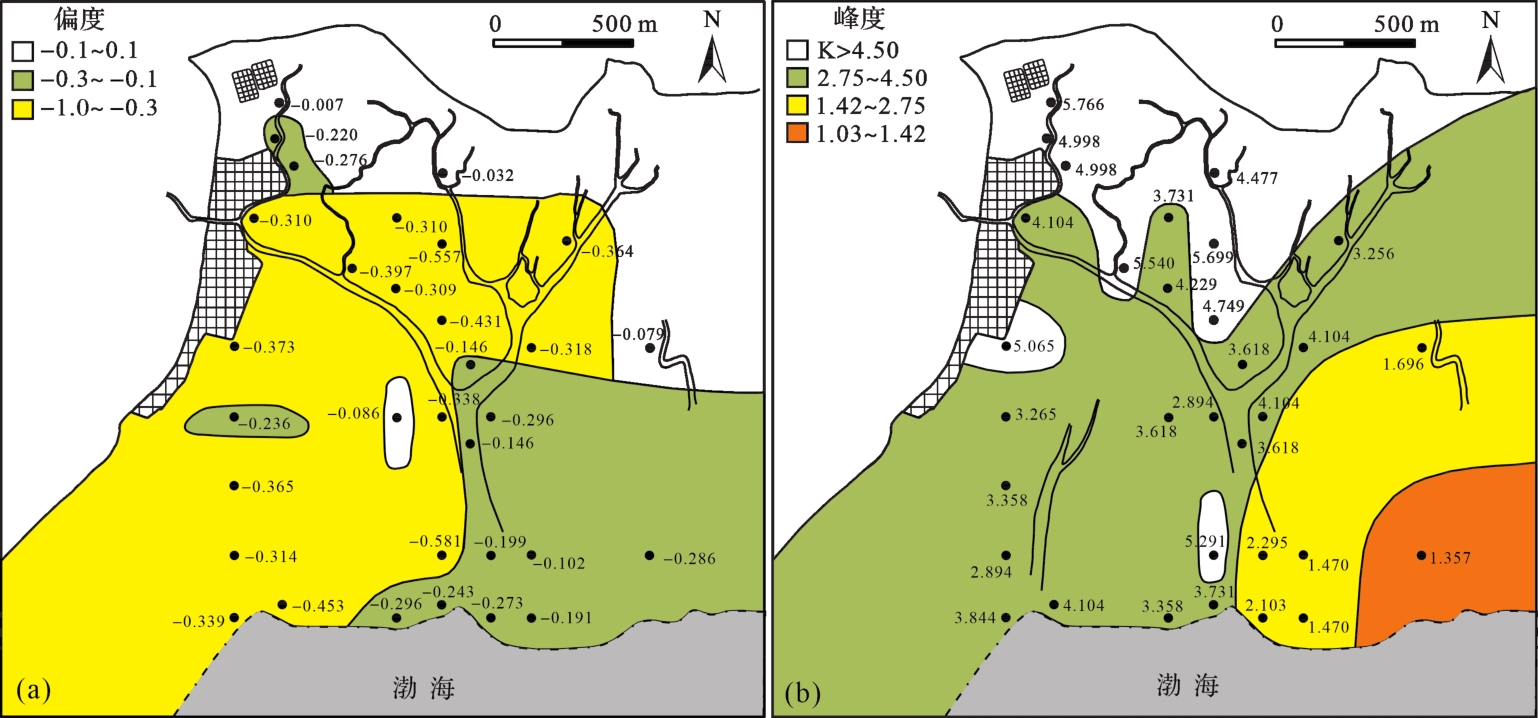
沉积物偏态值介于-0.60~-0.05,按照传统分类方式[43⁃44],由图6a可以看出,黄河三角洲潮坪内沉积物呈负偏或近对称。潮上带盐碱地的沉积介质速度比较稳定,粒度累计分布曲线大多是单峰近对称曲线,沉积物分选极好,偏态近乎对称(偏态值:-0.1~0.1)。潮道两侧和潮间带的采样点沉积物受潮水反复多次冲刷,在原先分选能力较强的沉积环境中得到了很好的分选,再搬运与潮道两侧沉积物发生混合,因此粒度累计分布曲线大多是单峰不对称曲线,偏度呈很负偏态(偏态值:-1.0~-0.3)和负偏态。受海水影响强烈的潮下带,粒度累计分布曲线大多呈尖锐双峰不对称曲线,偏度呈负偏态(偏态值:-0.3~-0.1),表明有新加入的沉积物。

Figure 6. Skew and kurtosis of sediments
粒度参数中峰度是用来衡量频率曲线峰凸程度。依据粒度累计分布曲线图中峰的个数来划分,潮上带和潮间带为单峰态,曲线偏斜程度高,因此峰态也高,峰值最大的部分居于中央峰,但其两侧有相对较宽的粗的和细的尾部,说明沉积物主要来自同一主体,来源相似且单一。潮下带为双峰态,沉积物粒径主要分布在3~4 Φ的极细砂组分,含有少量的6~10 Φ的细粒组分,分选相对潮上带和潮间带较差。按照McManus法划分标准[43],研究区峰度大于4.50占总数的51.5%,峰度2.75~4.50占总数的27.3%,峰度1.42~2.75占总数的15.2%,峰度1.03~1.42的占总数的3%,显示该区域的沉积物的峰态从非常平坦到相对尖锐所占比例逐渐减小。由图6b可以看出,从潮上带到潮下带峰度值逐渐变小,研究区最北部峰度值最高,达5.766,指示该区域表层沉积物混杂程度相对较低;在潮下带水动力强的区域,峰度值较低,表明该部分地区表层沉积物的混杂程度较高。
-
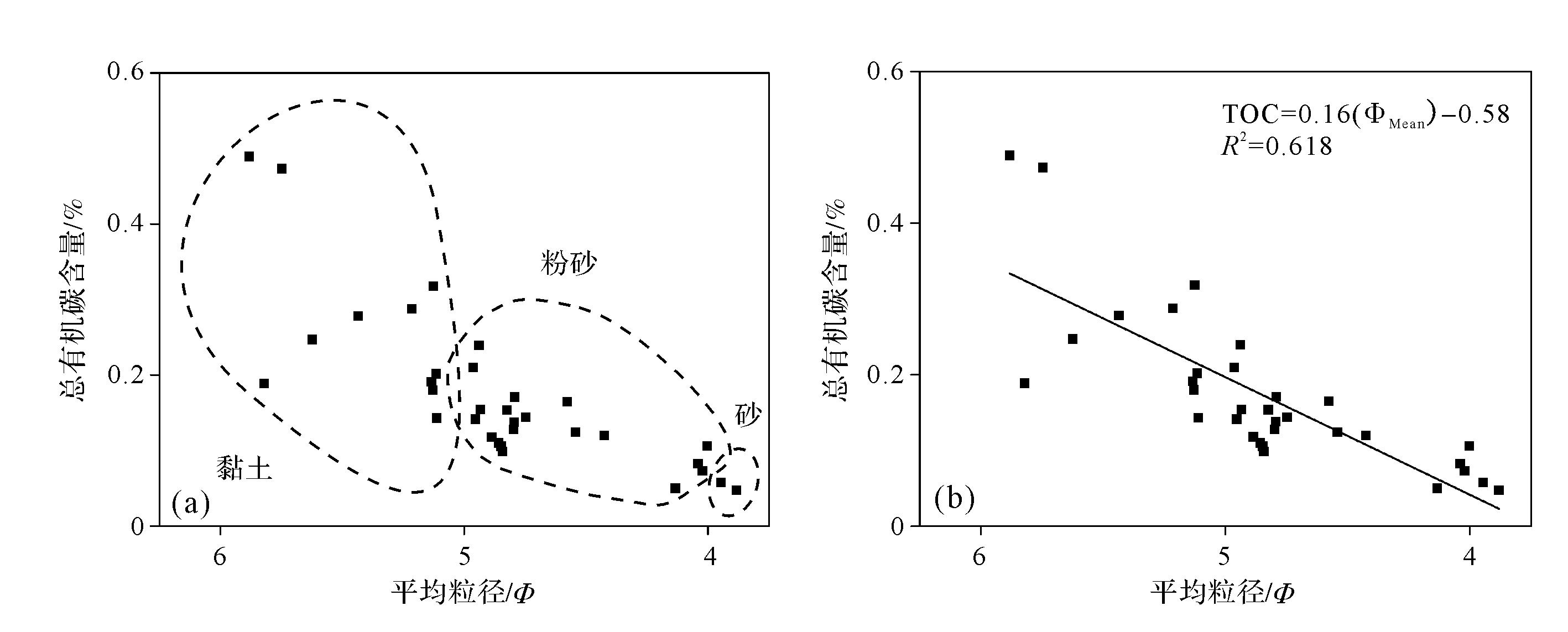
研究区大部分生物以沉积物和水中悬浮分散的有机物为食,即沉积物的总有机碳(TOC)含量[23]。当用phi值绘制TOC与平均粒径的关系时,很明显自潮上带到潮下带TOC随着平均粒径(ΦMean)的增加而减小,实际上,这种关系可以用公式(图7b)表示:
(1) 
Figure 7. (a) Relationship between total organic carbon (TOC) and mean grain size; (b) linear fitting of correlation between TOC and mean grain size
该方程定义较显著(R2=0.618),反映了TOC与粒度呈显著正相关,表明粒度是影响TOC含量的一个重要因素。潮上带不仅有潮汐带来的有机物,还有发育的植被“红地毯”,有机物含量高(图7a)。潮间带和潮下带水动力强粒度大,沉积环境相对复杂,受潮流等因素的影响,有机碳含量相对较低。
-
潮上带底栖生物种类相对较多,主要的造迹生物有甲壳类生物日本大眼蟹和豆形拳蟹;腹足类生物红带织纹螺、泥螺;蠕虫状生物双齿围沙蚕以及脊椎动物鸟。
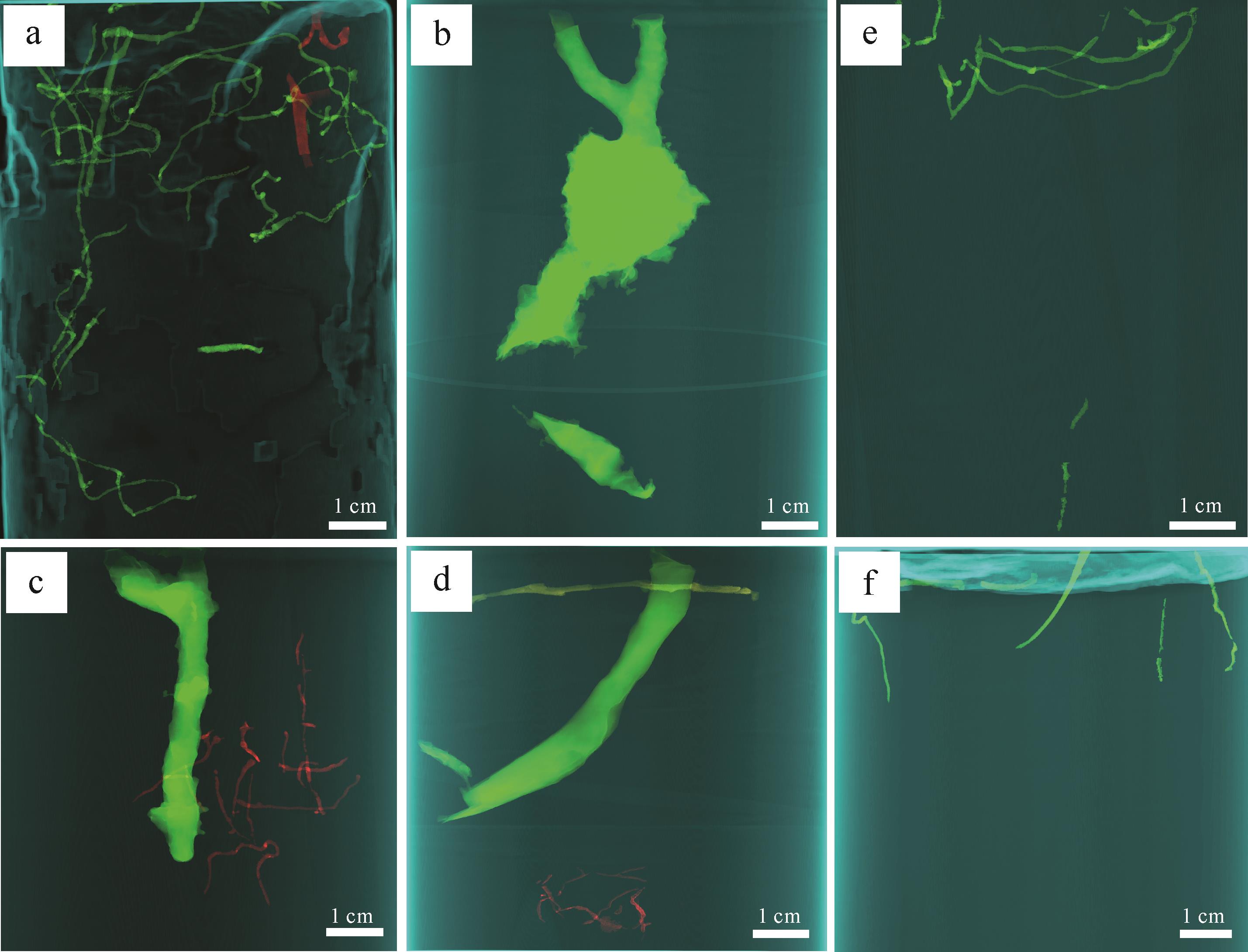
日本大眼蟹(Macrophthalmus japonicus)主要栖息在泥质基底上层10 cm深,扰动率为1.95%(表1),其生物遗迹主要为足辙迹、居住迹和排泄迹,爬行迹呈同心圆放射状、线性放射状和U型带状。在盐碱地的蛇曲形潮沟两侧,各点能量变化大,水动力较强,有机物来源丰富,生物扰动程度高,但分异度低,主要生物遗迹是日本大眼蟹的居住迹和足辙迹,其层面上爬行迹较少,形状呈放射状。潜穴口多为围墙状或漏斗状(图8a),下陷类似气坑构造,直径介于2~3 cm,形态多为Y型潜穴(图9b),分叉处有瘤状突起以加固潜穴,颜色与表层沉积物大不相同。在水动力强的潮道两侧,生物扰动强烈,但潜穴的保存是重要问题,因此挖掘了垂直或倾斜的潜穴,潜穴口呈土丘状(图8c)、鸟尾状,内部构造复杂,有Y型(图9c)、I型(图9d)、L型、U型和多分枝型(图9a)潜穴,在环境适宜的区域,弯曲型的柱体可延伸较深。
环境 生物 平均潜穴直径/cm 平均潜穴横截面积/cm2 占据点比例/% 平均潜穴密度/ (潜穴数/m2) 生物扰动/cm2 扰动率/% 扰动率(微环境)/% 潮上带 日本大眼蟹 2.19 4.22 28.13 41 195.41 1.95 5.05 豆形拳蟹 1.40 1.54 3.13 6 9.23 0.01 红带织纹螺 0.48 0.18 6.25 8 1.35 0.97 泥螺 0.45 0.17 6.25 569 97.32 0.63 双齿围沙蚕 0.42 0.15 12.50 405 63.21 0.09 鸟 4.52 16.22 9.38 9 139.38 1.39 潮间带 日本大眼蟹 3.41 9.73 12.50 31 329.35 3.29 2 946.88 红带织纹螺 0.53 0.22 6.25 23 4.98 0.05 泥螺 0.65 0.34 9.38 249 89.96 0.90 托氏昌螺 0.82 0.53 3.13 328 173.13 1.73 秀丽织纹螺 0.49 0.19 3.13 720 135.70 1.36 双齿围沙蚕 0.37 0.11 3.13 1 450 155.83 1.56 四角蛤蜊 2.05 3.87 6.25 15 65.63 0.66 牡蛎 13.55 144.13 3.13 2 037 293 588.66 2 935.89 鸟 3.40 9.07 3.13 16 145.19 1.45 潮下带 泥螺 0.82 0.53 3.13 254 134.07 1.34 22.99 秀丽织纹螺 0.63 0.31 3.13 42 13.09 0.13 托氏昌螺 1.49 1.74 3.13 1 048 1 826.43 18.26 四角蛤蜊 1.80 2.54 3.13 128 325.56 3.26 
Figure 8. The overview of a biodisturbance trace in the tidal flat of the Yellow River Delta

Figure 9. X⁃ray computed tomography images of burrows
豆形拳蟹(Philyra pisum de Haan)仅分布在研究区潮道边的泥质沉积底层中,扰动率最低,为0.01%。主要形成足辙迹和居住迹。沉积物含水率高,层面痕迹模糊不清,潜穴口形态单一,呈鸟尾状,与层面斜交,居住迹一侧有零散的沙球,或呈同心圆状的泥质突起,可能是生物进出时堆积而成。潜穴内部主要形态为Y型、U型和J型,且有双齿围沙蚕与其共生。
红带织纹螺(Nassarius succinctus)分布在研究区含水率高的泥坪中,爬行迹在沉积物表层呈弯曲的犁沟状。层内3D潜穴形态大多是I型,且扰动较深,也有Y型、U型,扰动率为0.97%。
泥螺(Bullacta exarata)分布于潮上带泥沙底,对盐度适应性强,潜于泥沙表层1~3 cm处,扰动率为0.63%。退潮后潮坪上泥螺大都浮出表面,拖迹形态呈极浅的条带状,中部平缓,两侧有略高的沙脊,其层面痕迹一般产生于含水量高的泥质汤底。
双齿围沙蚕(Perinereis aibuhiteusis)造迹主要是拖迹、觅食迹和居住迹。研究区观察到的层面拖迹模糊不清,星射状的觅食迹与潜穴口相连(图8b),生物扰动率仅为0.09%。潜穴形态复杂多样,有Y型、U型及复杂分枝状,分叉处有囊泡状突起,可以与螃蟹共生(图9c),也可以独自掘穴(图9d),扰动深度通常为8~20 cm。
鸟足迹在中高潮区居多,是由于研究区的中高潮区生长植被以及大量的泥螺,为鸟类提供了丰富的食物,因此足迹清晰且量多。潮上带的扰动率为1.39%,潮间带的扰动率是1.45%,扰动程度相近。
总体来说,潮上带生物总扰动程度最低,扰动率为5.05%。潮上带盐碱地丰度值相对较高,生物遗迹分异度最低。
-
潮间带底栖生物种类最多,主要的造迹生物有甲壳类生物日本大眼蟹;腹足类生物红带织纹螺、泥螺、托氏昌螺、秀丽织纹螺;蠕虫状生物双齿围沙蚕;双壳类生物四角蛤蜊和牡蛎,以及脊椎动物鸟。
日本大眼蟹在沙泥质沉积物表层及内部活动,生物遗迹主要为足辙迹和居住迹。由于沉积物含水量以及粒度的不同,其层面痕迹和潜穴口形态不一。爬行迹呈线性放射状,足辙迹呈U型带状,在含水率高的沉积物表层,痕迹模糊不清,含水量适中时痕迹清晰。潜穴口呈下陷椭圆状、鸟尾状,形态主要为Y型、U型、L型和I型,尤其在主潮道两侧的潜穴重构中发现其内部构造复杂,有瘤状突起,涨潮时可以躲居在潜穴中,生物扰动较潮上带高,扰动率为3.29%。
红带织纹螺分布在研究区含水率高的混合坪中,主要为U型带状爬行迹(图8f),扰动率为0.05%,较潮上带低,与螃蟹共生,层内潜穴形态大多是I型、Y型、U型及复杂分支型(图9e)。
泥螺栖息在潮间带低潮区,扰动率为0.9%,形成的遗迹主要为拖迹,形态不一,有直型、任意弯曲型,不分叉,但是由于沉积物为过饱和的软底,因此痕迹模糊不清。
托氏昌螺(Umbonium thomasi)生活在砂泥混合坪中低洼有水的地方,生物扰动率为1.73%,主要为U型条带状的运动迹,呈现出规则的羽状纹理,中间有极细的突起沙脊,两侧纹理对称且与沙脊成一定角度。
秀丽织纹螺(Nassaurius festiva)分布于研究区的混合坪中,沉积物含水率高,生物潜在表层浮沙下运动,仅可见模糊不清的弯曲带状痕迹,具体细节无法观察,扰动率为1.36%,生物扰动程度较潮下带(0.13%)高。
双齿围沙蚕层面上可见U型条带状不分枝的运动拖迹,以及沿潜穴口大致呈星射状分布的觅食拖迹。层内垂直管状,有J型、Y型、U型的居住迹和觅食潜穴,穴道为圆形,内表面光滑。由于潮间带低洼积水处、潮道两侧沉积物颗粒较细,含水量高,有利于沙蚕向下挖掘生存,因此扰动率较潮上带高出许多,扰动率为1.56%。
牡蛎(Ostrea gigas tnunb)多分布在半咸水环境,在浅层泥沙中穴居(图8e),丰度极高,生物扰动率为2 935.89%。进食方式以过滤为主,依靠海洋中的微型海藻和有机碎屑为食。
四角蛤蜊(Mactra veneriformis Reeve)呈卵圆形,通过虹吸管进食,主要为层内穴居,潜穴与层面近乎于垂直,三维重构可见层内有与生物外形一致的印痕。
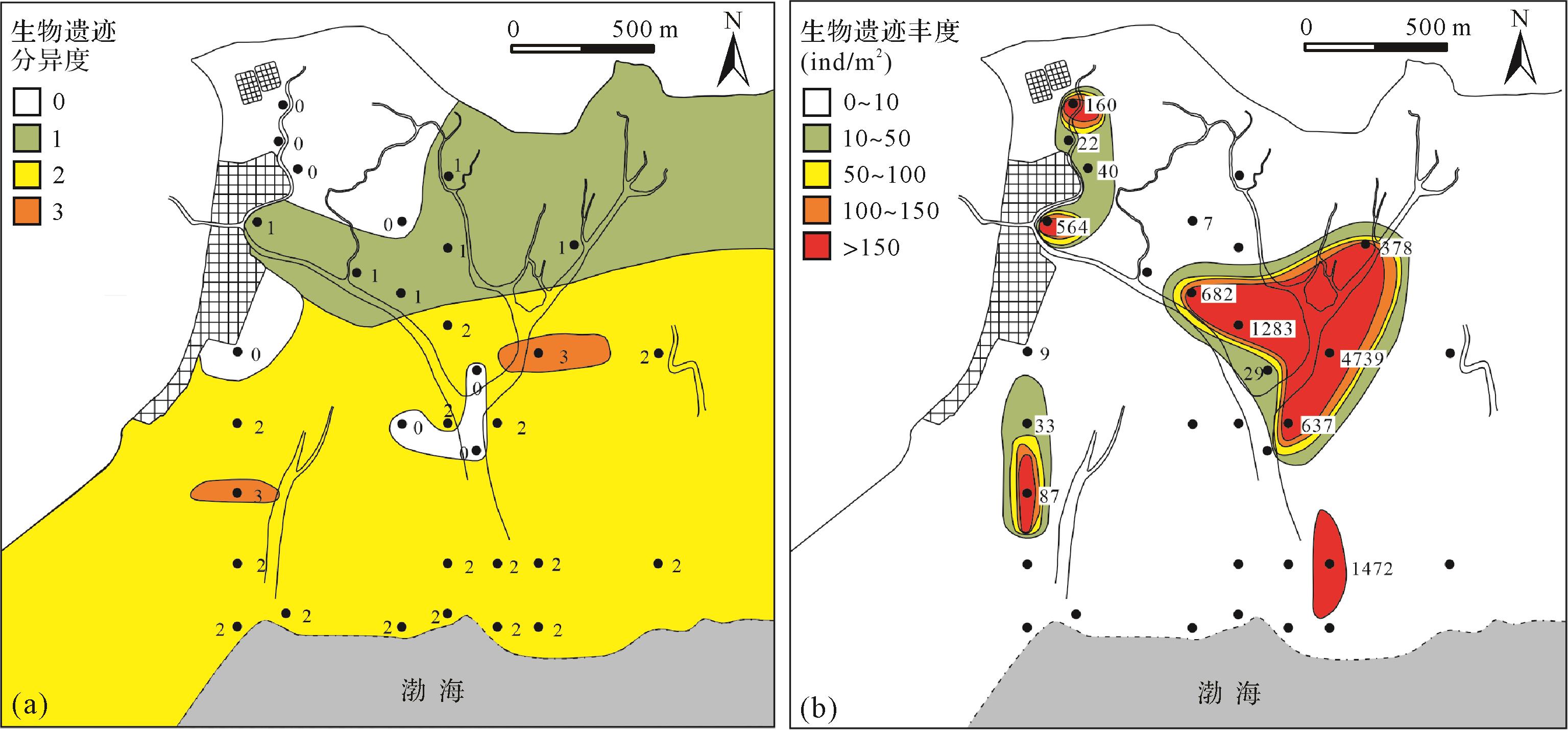
潮间带生物扰动程度最高,扰动率为2 946.88%,主要是因为潮道口牡蛎扰动程度非常高,如果减去牡蛎的扰动率,其他生物扰动率总和为10.99%,那么相较潮上带和潮下带,其生物扰动程度将次于潮下带。潮道两侧潮汐往复,丰度值较潮上带逐渐增大,并在潮间带潮道口处达到最大值4 739 ind/m2,分异度也达到最大值3。
-
潮下带生物遗迹的观察和取样于低潮期乘船时进行,泥螺和秀丽织纹螺的生物遗迹特征与潮间带相似。托氏昌螺在沉积物表面丰度极高,且爬行迹杂乱无章(图8d),而四角蛤蜊密集潜居沉积物下3~10 cm处,大多呈相同方向排列。通过对托氏昌螺的重构,发现其形态与沉积物表层呈倾斜状(图9f),掘穴深度相对较浅。潮下带的生物扰动率为22.99%,丰度值较潮间带降低,而分异度基本保持不变。
3.1. 物理沉积构造
3.2. 沉积特征分析
3.3. TOC特征分析
3.4. 生物遗迹特征
3.4.1. 潮上带生物遗迹
3.4.2. 潮间带生物遗迹
3.4.3. 潮下带生物遗迹
-
整体来看,黄河三角洲潮坪生物遗迹在空间分布上存在不均衡性及其复杂性,潮间带的生物扰动程度最高(图10),潮下带次之,潮上带最低。而生物遗迹的丰度和分异度平行于潮坪的地形地貌特征,呈现出从潮上带到潮下带逐渐增大的趋势,在潮间带潮道汇聚处达到最大(图11)。但是其中的生物遗迹分布特征的精细差别与潮坪环境的物理化学条件具有良好的响应关系。主要的物理化学条件包括:浑浊度、盐度、水动力特征、粒度、沉积速率及TOC。

Figure 10. The distribution of bioturbation

Figure 11. The heterogeneity and abundance of biological traces
-
水体的浑浊度会对生物遗迹的种类产生影响,如高浑浊度的水阻碍了食悬浮物造迹生物的滤食行为,堵塞造迹生物的虹吸管[45⁃47]。尤其对于高浑浊度河流影响下的黄河三角洲潮坪来说,浑浊度将对生物遗迹的种类产生重要的影响[48]。浑浊度平均值为287.02 NTU(NTU是散射浊度单位),潮上带粒度和水动力均小,因此浊度值最小32.40 NTU,浑浊度对生物遗迹的丰度和分异度基本没有影响。随着潮道的加宽,水深变深,潮流的剧烈冲刷,浊度逐渐上升且普遍高,从而呈现出潮上带到潮下带增加的趋势(图12a),在潮道拓宽处达到最大值936.40 NTU,在多条潮道汇集处浑浊度降低,从而生物遗迹的分异度、丰度和扰动率呈先增加后减小的趋势(图13)。这是由于潮间带主要造迹生物如甲壳类生物日本大眼蟹、豆形拳蟹和软体动物红带织纹螺、泥螺、托氏昌螺、秀丽织纹螺等以泥质有机物为食物[49],由于浑浊度的升高,食物来源匮乏,生物遗迹丰度和分异度急剧减少(表1)。而在多条潮道汇集处由于牡蛎礁的缓冲作用,浑浊度降低,以食悬浮物为主的双齿围沙蚕、双壳类生物四角蛤蜊等造迹生物虹吸管的滤食行为大大增强,该类生物数量增多(表1),扰动率增强。而潮下带在潮汐涨潮期间堆积的泥沙流增加水浊度,生物遗迹的分异度不变、丰度略微降低(图13)。这是由于潮下带的造迹生物主要为托氏昌螺和四角蛤蜊,增大的浑浊度大大限制了这些食悬浮物的滤食效率,因此生物遗迹丰度略微降低。

Figure 12. Salinity and turbidity

Figure 13. The influence factor mode of the neoichnology assemblage in tidal flat of the Yellow River Delta
-
生物对盐度的敏感性表现在物种组成、生物现存量、多样性存在明显的空间不均衡性。盐度的增加和减少以及生物耐盐性,也是影响生物群落的分布的关键因素[50⁃51],这是因为盐度可以显著影响生物群落中喜盐生物和抗盐生物的分布。随着涨潮,会流入低盐度水域,底栖动物的物种丰度明显减少,因此盐度对潮坪遗迹的分异度、丰度、扰动程度、遗迹的直径、遗迹的分布特征及分布形式等均具有灵敏的影响[52⁃55]。黄河三角洲潮坪盐度特征整体与地形地貌特征平行,但是受潮道分布的影响,潮坪盐度特征在横向上呈差异性分布。生物遗迹与盐度特征响应关系明显。
研究区长期受海水侵蚀,盐分较高,属于半咸水和咸水持续混合区域(图12b),潮上带靠近农田区域盐度相对较低,属于半咸水环境(0.5‰~16‰),整体生物扰动率中等,生物遗迹的丰度和分异度中等(图13)。在远离潮道的区域,盐度低,生物遗迹丰度高,但是生物遗迹的分异度与潮上带大体持平。日本大眼蟹属于广盐性生物[56],在盐度过高的情况下不适应。当达到最适盐度26.25‰时,生物丰度最大,生物扰动程度也最高,平均潜穴直径为2.19 cm,扰动密度41个/m2。潮间带盐度稳定,生物扰动率、生物遗迹的丰度和分异度最高,尤其在潮道入海口处,盐度降低,生物扰动率达到最高值,生物遗迹的丰度达到最高值,同样的日本大眼蟹的潜穴直径增加到3.41 cm,但是扰动密度减至31个/m2,这是由于盐度的降低导致日本大眼蟹数量及其潜穴数量的减少,螃蟹为了适应盐度的降低不得不增加挖穴的直径以获取更大量的咸水,调整潜穴内的盐度。潮下带盐度高,属于咸水环境(16‰~47‰),高营养盐和有机碎屑随着潮汐进入潮下带,盐度整体升高,海洋生物比例大于淡水生物,生物遗迹的丰度和分异度中等,主要造迹生物为喜盐性生物托氏昌螺和四角蛤蜊[57]。
-
水动力、粒度及沉积速率三者之间是相互联系的,水动力条件和粒度对沉积速率起到了关键的控制作用[24],而TOC又与粒度特征呈正相关(图7),四者共同影响了生物的分布特征。水动力主要来自波浪和潮汐,基本没有受到黄河的直接影响,其中波浪具有明显的季节性,以冬季最甚,春季和秋季次之,夏季最弱,潮流为不规则的半日潮,潮汐携带黄河扩散至海洋的泥沙进入潮坪,因此沉积物主要为黄河入海泥沙和再悬浮沉积物[31,38]。
潮上带为低能高潮区,以泥质沉积物为主,极个别采样点为粉沙,分选极好,粒度累计分布曲线大多是单峰近对称曲线,偏态近乎对称,峰态非常平坦,水动力弱,有机物来源丰富,TOC含量最高,生物遗迹分异度最低。然而在粉沙质沉积物区域生物遗迹丰度最高(图13),这是因为该区域的底质含水量较小、较坚硬,适合日本大眼蟹生存[49]。潮道两侧日本大眼蟹的丰度较其他生物高,因为他们更能适应复杂多变的水文和沉积环境[58]。
潮间带沉积物为粉沙,分选极好,粒度累计分布曲线大多为单峰不对称曲线,偏态呈很负偏,峰态平坦,坡度平缓,生物遗迹分异度和丰度整体增高。在几条潮道汇聚处往复的潮汐带来的有机物又为动物提供了充足的营养物质,TOC含量相对较高。水动力相对潮下带较弱,沉积速率低,生物遗迹丰度和分异度达到最高,尤其是潮道口的大片牡蛎礁,对湍急的风浪起到一定的缓冲作用,为其他生物创造了较好的生存环境[59]。
潮下带属于高能低潮区,以极细砂为主,分选极好,粒度累计分布曲线大多呈尖锐双峰不对称曲线,偏态呈负偏,峰态相对尖锐。物理沉积构造相对简单,粒度最粗、TOC含量最低,水动力最强,沉积速率最高,生物遗迹分异度基本保持不变,丰度增高,造迹生物种类变化,主要为托氏昌螺的爬行迹和四角蛤蜊的居住迹。
-
首先,在古潮坪遗迹学方面,黄河三角洲潮坪不能简单地用任何一类已建立的遗迹相来描述。黄河三角洲潮坪环境的生物遗迹既有层面类型又有层内类型,混合了典型的Skolithos遗迹相和Cruziana遗迹相。一种新的适用于黄河三角洲潮坪在物理化学环境因素影响下的类古潮坪环境的遗迹相模型有待建立。
其次,在古潮坪沉积环境方面,黄河三角洲潮坪的现代生物遗迹为类比研究古潮坪的沉积速率提供了现代遗迹方面的实证材料。三角洲潮坪的低沉积速率有利于造迹生物在沉积底层造迹以及生物遗迹的更均衡分布和保存,而高沉积速率条件下,底层沉积物间歇性瞬时性的迅速沉积导致生物生殖、居住活动锐减,生物遗迹丰度随之降低。另外,随着沉积物的迁移和瞬时沉积,动荡和不稳定的环境限制了造迹生物的种类,从而导致生物遗迹分异度降低。
4.1. 浑浊度与生物遗迹分布的关系
4.2. 盐度与生物遗迹分布的关系
4.3. 水动力、粒度、沉积速率、TOC与生物遗迹分布的关系
4.4. 对古潮坪遗迹学和古潮坪环境的启示
-
(1) 生物遗迹的分异度、丰度所受到的环境压力主要是盐度降低、沉积速率增加、水浊度增加、水动力强弱、沉积物的粒度大小和TOC含量,其中盐度和浑浊度影响了生物遗迹分布的种类和数量。造迹生物种类相同的情况下,盐度影响了生物潜穴的直径和扰动率;盐度相同的情况下,高浑浊度限制初级生产力,影响生物潜穴的数量。
(2) 潮上带生物遗迹分异度在盐碱地最低,仅有日本大眼蟹的爬行迹和居住迹。在粉砂质底层的区域和多条潮道的汇聚处,生物遗迹的丰度和分异度最高,主要为甲壳类生物日本大眼蟹的足辙迹、居住迹和排泄迹,豆形拳蟹的足辙迹和居住迹;腹足类生物红带织纹螺的爬行迹和居住迹,泥螺的爬行迹;蠕虫状生物双齿围沙蚕的拖迹、居住迹和觅食迹,以及脊椎动物鸟足迹。
(3) 潮间带整体生物遗迹分异度和丰度较潮上带和潮下带高,但在TOC含量相对较高、水动力相对较弱、沉积速率低、盐度降低的几条潮道汇聚处生物遗迹丰度和分异度达到最高,主要为甲壳类生物日本大眼蟹的爬行迹、居住迹和排泄迹;腹足类生物红带织纹螺的爬行迹和居住迹,泥螺的爬行迹,托氏昌螺的足辙迹,秀丽织纹螺的爬行迹;蠕虫状生物双齿围沙蚕的拖迹、居住迹和觅食迹;双壳类生物四角蛤蜊和牡蛎的居住迹,以及脊椎动物鸟足迹。
(4) 潮下带生物遗迹丰度和分异度整体呈对称均衡分布,主要为托氏昌螺的爬行迹和四角蛤蜊的居住迹。

 DownLoad:
DownLoad: