

 下载:
下载:



-
鲕粒是碳酸盐岩中重要的颗粒类型,形态通常为直径小于2 mm的、有核心的、被均匀圈层结构包裹的球型或椭球形,可以用来指示古环境条件(水深、盐度、温度和水体能量)[1⁃6],由鲕粒构成的鲕粒石灰岩也是碳酸盐岩中优势的储集层[7⁃9]。鲕粒在形态上与核形石相似,但核形石生物成因被普遍接受[10⁃11],而自Sorby[12]提出鲕粒以“滚雪球”的方式生长以来,关于鲕粒的物理化学成因还是生物成因就一直存在争论。争论主要集中在:1)微生物是否在鲕粒圈层形成中起到积极作用;2)光合微生物和硫酸盐还原细菌微生物群落在鲕粒形成中是否发挥作用;3)鲕粒的放射状结构是次生的还是原生;4)鲕粒是否是海平面变化的可靠指标;5)直径大于2 mm甚至达到厘米级的巨鲕在深时地层中也有发育,这种鲕粒与传统粒径较小的鲕粒在成因上有没有差异。现今正在沉积的海相鲕粒普遍发育在高盐度、高碱度、碳酸钙过饱和的浅水环境中[13],而日内瓦淡水湖泊鲕粒的发现似乎表明高碳酸盐饱和度的水体不再是鲕粒形成的必要条件,相反微生物作用发挥了更重要的作用[14⁃15]。
本文系统归纳总结了鲕粒成因的主要观点及鲕粒圈层结构类型,在综合各种观点的基础上,提出了一种可能的鲕粒形成模式,并探讨了鲕粒在沉积环境判别方面的指示意义。
-
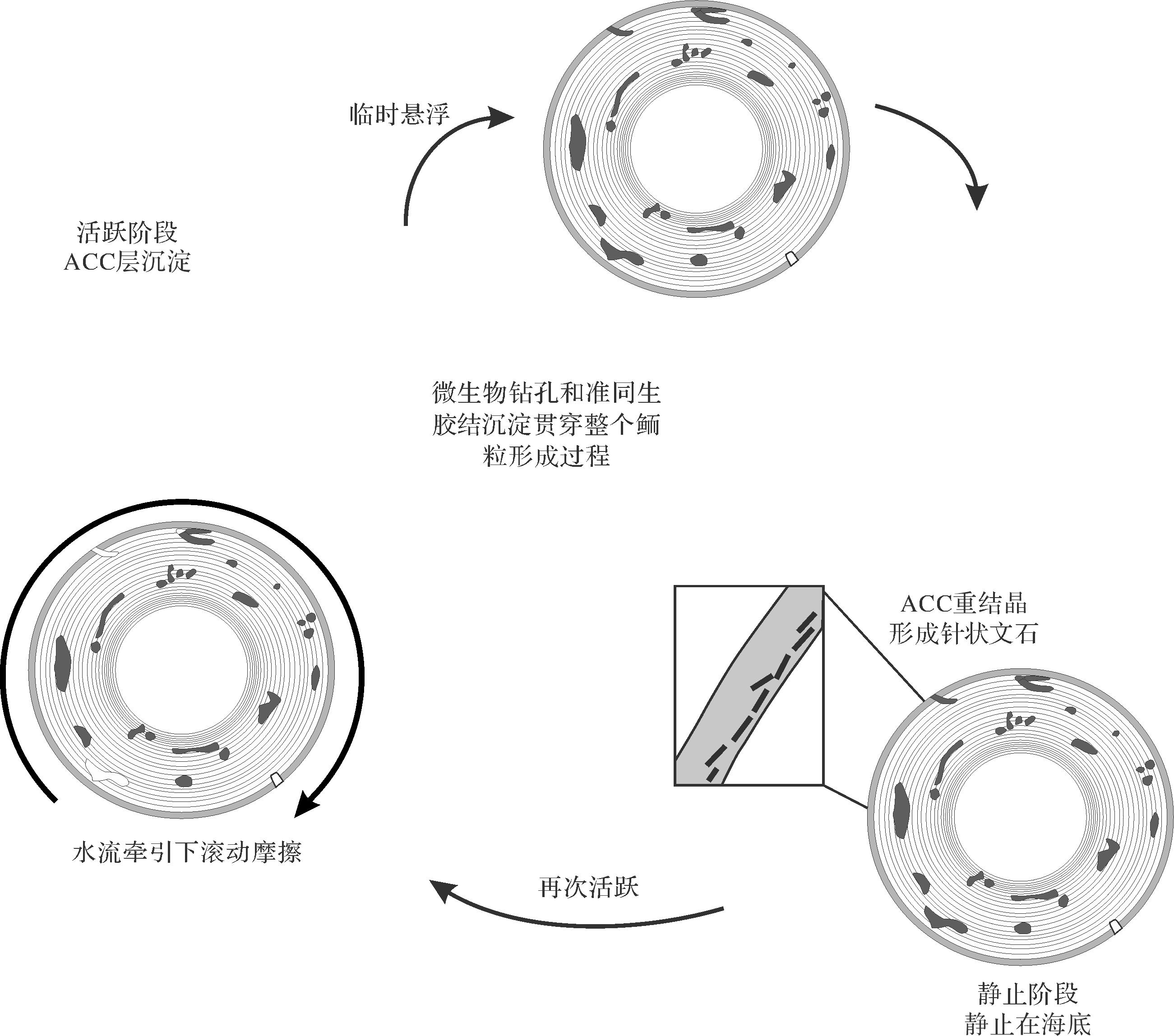
典型鲕粒的物理化学成因假说认为鲕粒形成需要三个条件:1)碳酸盐过饱和的海水;2)作为核心的颗粒来源;3)动荡的水体。Davies et al.[16]通过实验模拟在安静的水体中合成了径向生长的放射鲕,在动荡的水体中合成了切线生长的同心鲕,进而认为鲕粒是无机沉淀形成的;认为活跃阶段鲕粒表面Mg2+的聚集“毒化”了鲕粒,使鲕粒停止生长,而静止阶段通过去除Mg2+使鲕粒表面重新活化,从而周期性产生围绕核心的叠片状同心层。鲕粒形成所需要的物理化学地质条件在现代热带碳酸盐浅滩是比较普遍的,但不是所有的浅滩都有鲕粒的发育。最近的全球碳酸钙地球化学研究表明海洋中存在一定的pH值和碱度的变化[17]。Rankey et al.[13]通过对太平洋库克群岛现代鲕粒研究发现其主要形成于群岛附近pH值和碱度最高的地区,其他地区鲕粒基本不发育,并且现代发生鲕粒沉积的海域具有相同的特征——高pH值和高碱度,在适当的水动力环境下,有利于提高碳酸钙过饱和状态,促进鲕粒的形成,认为碳酸钙过饱和状态是限制鲕粒全球分布的重要因素。Duguid et al.[18]研究认为微生物在鲕粒的形成中没有起到建设性作用,相反,由于微生物的存在造成钻孔及与其相关胶结物的形成,改变了原始鲕粒圈层的结构和化学组成,并将鲕粒形成划分为两个阶段(图1):1)活跃阶段,鲕粒表面形成具有相对较高的Mg/Ca比的非晶质钙碳酸盐ACC(Amorphous Calcium Carbonate);2)稳定阶段,ACC重结晶形成针状文石,组成鲕粒新圈层(图1)。鲕粒外层Mg2+浓度的升高与ACC的形成密切相关,是新的鲕粒圈层的形成而不是抑制鲕粒生长[18],据此可以推断Davies et al.[16]认为的Mg2+“毒化”鲕粒的过程其实是形成ACC的过程。鲕粒圈层中有大量的微生物遗迹,这些微生物及钻孔普遍是鲕粒在稳定阶段被光合作用微生物栖息的结果[18],Trower et al.[19]的研究也表明鲕粒的生长主要受碳酸盐饱和度和运移模式的影响,微生物在鲕粒表面通过微孔化破坏了鲕粒原始结构。

鲕粒的物理化学成因表明鲕粒极有可能是以围绕核心周期性的ACC膜为模板钙化形成鲕粒圈层,高pH值、高碱度和核心的来源是海相鲕粒形成必不可少的条件。虽然现代和古代鲕粒中均存在大量的微生物痕迹,但都是微生物定殖在鲕粒外表的结果。虽然有学者在实验室合成鲕粒并认为现在巴哈马鲕粒形成是无机过程[16],但实验却不是在完全没有有机物质的参与下进行的,并且鲕粒为了进一步生长,需要有机膜提供基质。Duguid et al.[18]虽然提出了以ACC为模板的鲕粒形成模式,但却没有解释清楚ACC是如何形成的。
-
无论是深时地层中的鲕粒,还是现在正在沉积的水体中的鲕粒,都发现了微生物痕迹,因此产生了微生物介导碳酸钙沉淀的鲕粒微生物成因观点。
-
Folk et al.[20]研究认为大量鲕粒的沉淀与纳米细菌有关,并且鲕粒中文石大致切向的取向是由于黏液的“定向模板”造成的,而不是随机的。黏液填充的层与文石层周期性排列构成了鲕粒的圈层结构,而这些富有机质的黏液层在后期腐烂形成了文石层之间的间隙。Summons et al.[21]研究认为特定的微生物群落定殖在鲕粒圈层表面,并可能促进鲕粒皮层的生长,强调了硫酸盐还原细菌对鲕粒形成的重要性,海洋底栖微生物群落中硫酸盐还原菌的存在和代谢活性增加了局部的碱度,促进了碳酸盐的沉淀,这些特定的微生物在鲕粒形成中活跃,控制了鲕粒的钙化。Tang et al.[22]通过对华北雾迷山组的研究认为,鲕粒中大量残余的有机质证明微生物在鲕粒的形成中起到了至关重要的作用,细菌丝通过原位矿化形成鲕粒,细菌丝束在鲕粒生长方向上的保存需要在碳酸钙过饱和环境中快速矿化。Tan et al.[23]在早寒武世鲕粒中发现了大量生物遗迹,从而说明微生物在鲕粒形成中发挥了重要的作用,并且放射鲕形成是微生物和高饱和度碳酸钙共同作用,与Tang et al.[22]研究结论相似。Diaz et al.[24-26]研究表明微生物群落可以诱导鲕粒圈层的形成,由细菌活动诱导的碱度和过饱和条件的局部增加可导致ACC发展和随后的文石晶体形成,强调了ACC形成的生物过程;鲕粒是由微生物介导的有机矿化作用形成的,ACC在鲕粒中的文石晶体发展中起到作用,与经典的成核理论相反,通过ACC的预核化发生鲕粒矿化,其中ACC与EPS和微生物之间的密切关联表明ACC沉淀物是生物介导的;将鲕粒的形成总结为以下四个阶段(图2):1)鲕粒外部圈层EPS形成导致碳酸钙的沉淀形成ACC;2)ACC的累积形成薄层;3)ACC转化为文石晶体,形成围绕核心的新的文石层;4)动荡的水体磨损不均匀的文石层,形成较为光滑的球形,构成了鲕粒的圈层(图2)。除此之外,也有众多学者支持鲕粒的微生物假说[27⁃28]。
-
瑞士的日内瓦湖淡水中发现了大量以石英碎屑和碳酸盐生物介壳为核心的同心鲕。原地实验表明光合微生物群落在鲕粒圈层的形成中起到了至关重要的作用,如丝状和球状的蓝藻,在相同的物理化学条件下,生物膜的缺乏抑制了方解石的沉淀[15]。淡水湖中光合微生物不仅促进了鲕粒核心周围早期碳酸盐岩的沉淀,而且控制了鲕粒整个圈层的形成,硫酸盐还原细菌对鲕粒的形成基本不起作用,鲕粒圈层在钙化之前首先由光合微生物细胞外聚合物质(EPS)矿化为无定型硅酸镁(amMg-Si),为鲕粒碳酸钙沉淀提供模板,然后转变为低镁方解石[14]。Brehm et al.[29⁃30]通过球形微生物群落在实验室合成了类似于鲕粒的同心结构,丝状蓝细菌的球形排列产生空心球,碳酸盐晶体沿着蓝细菌沉淀,认为这是一种鲕粒的前体,是由微生物群落主导形成的。
鲕粒中残余大量的有机质,鲕粒成因微生物假说表明微生物作用促进了碳酸盐沉淀,物理化学假说表明微生物的活动破坏了鲕粒圈层结构,这很可能与微生物的类型有关。以钻孔为主的微生物对鲕粒起到破坏作用,能增加局部碳酸钙饱和度从而促进碳酸钙沉淀的微生物等对鲕粒形成起到建设性作用。湖相淡水鲕粒支持了光合微生物(如蓝藻)对鲕粒圈层的建设性作用,Summons et al.[21]通过测定鲕粒内的生物标志表明光合作用微生物、异养微生物特别是硫酸盐还原细菌均有助于鲕粒的形成,Tang et al.[22]推测活跃的硫酸盐还原细菌是华北地区雾迷山组巨鲕形成的主要原因之一,Diaz et al.[24⁃26]与Tan et al.[23]也支持光合作用微生物及硫酸盐还原细菌等均可以改变局部碳酸钙饱和度和碱度,从而促进鲕粒圈层形成。因此,可以将对鲕粒有影响的微生物分为两类:一类是以光合作用微生物及硫酸盐还原细菌为主的促进鲕粒新圈层形成的建设性微生物群落;一类是以强烈的微钻孔为主的破坏鲕粒原始结构的破坏性微生物群落。
-
根据鲕粒圈层结构类型将鲕粒分为同心鲕、放射鲕、同心放射鲕、晶体鲕及泥晶鲕等。晶体鲕是受成岩作用改造重结晶形成的,泥晶鲕是在沉积时期或沉积后受生物活动的影响原始圈层结构遭到破坏形成的。虽然宋文天等[31]将鲕粒圈层类型分为两大类七小类,但与碳酸盐矿物相关的鲕粒圈层主要分为三类(图3):1)围绕核心径向展布的放射状结构;2)围绕核心切线状分布的同心状结构;3)围绕核心随机分布的杂乱结构(图3)。这三种结构既可以同时出现在一个鲕粒中,也可以单独存在一个鲕粒中。

图 3 鲕粒典型圈层结构展示
Figure 3. Characteristics of representative oolitic structure
现在的巴哈马鲕粒均为切线状同心鲕,因此同心鲕普遍被认为是原生的,而在放射鲕是原生和次生问题上存在争议。Rao et al.[32]通过对印度西北部的鲕粒研究认为径向取向的结构是次生的,形成于早成岩阶段安静的水体,与皮质中微生物薄层的矿化作用密切相关。Sandberg[33]通过对大盐湖鲕粒研究认为放射状径向取向的鲕粒结构是原生的,不是成岩作用形成的,其保留与有机物质密切相关。Siahi et al.[27]通过对南非Mesoarchaean Chobeni组鲕粒的研究认为放射鲕来源于低能带有微生物参与的高镁方解石,同心鲕是文石在动荡高能的环境中形成的,同心—放射鲕是由双矿物构成的,代表了海洋水体成分的变化。Davies et al.[16]通过实验模拟在有机质参与的低能环境中合成了放射状生长的鲕粒。
现今众多的证据支撑了鲕粒放射状结构是原生的,而无论是鲕粒的物理化学成因假说还是微生物成因假说,均支持放射状径向生长的鲕粒结构需要低能安静的水体环境,切线状同心结构是低能环境中形成的放射结构在高能环境中磨平改造的结果,因此鲕粒类型是判断水动力变化的可靠指标。
虽然鲕粒内部结构的差异性可以是不同类型矿物构成的,例如放射鲕普遍起源于高镁方解石,文石也可以形成放射结构,同心鲕普遍由文石构成[27,34],但是作者认为水动力条件是鲕粒不同结构形成的主要原因,严格意义上的切线状圈层是不存在的,鲕粒圈层由径向—随机—切线结构演化,代表了沉积水体能量的增加。周瑶琪等[35]通过实验模拟也表明扰动较小的安静水体有利于晶体垂直鲕粒表面纵向生长,水动力的存在对纵向生长的晶体产生了定向改造,使晶体沿切向排列,进一步说明了水动力改造是不同鲕粒圈层结构形成的主要原因。
-
鲕粒粒径通常小于2 mm,但不可忽视在地质历史时期还存在大于2 mm甚至达到厘米级别的巨鲕。Sumner et al.[36]研究表明鲕粒大小是反映沉积环境物理化学特征的重要标志,低核心供给速率、高碳酸钙饱和度的水体、高鲕粒圈层生长率和高平均水流速度及水流梯度有利于巨鲕的形成;Tang et al.[22]通过对华北雾迷山组鲕粒研究表明巨鲕主要发育在浅滩开阔水域较深水且搅动较弱的低氧至缺氧环境中,硫酸盐还原细菌的活跃导致碳酸钙浓度增加促进了巨鲕的形成;段雄等[37]研究表明频繁的强风暴背景和蓝细菌等微生物的积极参与是重庆石柱地区早寒武世巨鲕形成的必备条件;Trower et al.[38]研究表明鲕粒的粒径大小是一个圈层生长和磨损的动态平衡过程,而磨损率并不是随着动荡程度单调增加,反而强动荡的水体造成大部分鲕粒悬浮,延长了鲕粒的生长周期,导致更大鲕粒的形成。Li et al.[39]研究认为高温和干旱气候导致的高碳酸钙饱和度是华南早三叠世鲕粒大面积分布的主要原因,因而碳酸钙饱和度也是该时期大于2 mm鲕粒普遍发育的主要原因。
更高碳酸钙饱和度的水体及低的核心供给速率有利于更大粒径鲕粒的形成已经成为共识。虽然Tang et al.[22]认为巨鲕形成于较深水的安静环境,但相对动荡的环境有利于更大的鲕粒悬浮生长。相对更加动荡的水体有利于鲕粒周期性悬浮生长,从而形成更大粒径的鲕粒,这也是同一沉积环境中静水放射鲕粒径普遍小于动荡同心鲕的主要原因,而且动荡的水体降低了鲕粒在间歇期沉积的概率,提高了鲕粒的生长周期,有利于更大粒径鲕粒的形成。鲕粒新圈层的形成需要建设性微生物群落介导EPS和后续ACC的形成,活跃的建设性生物群落作用提高了鲕粒圈层的生长速率。
-
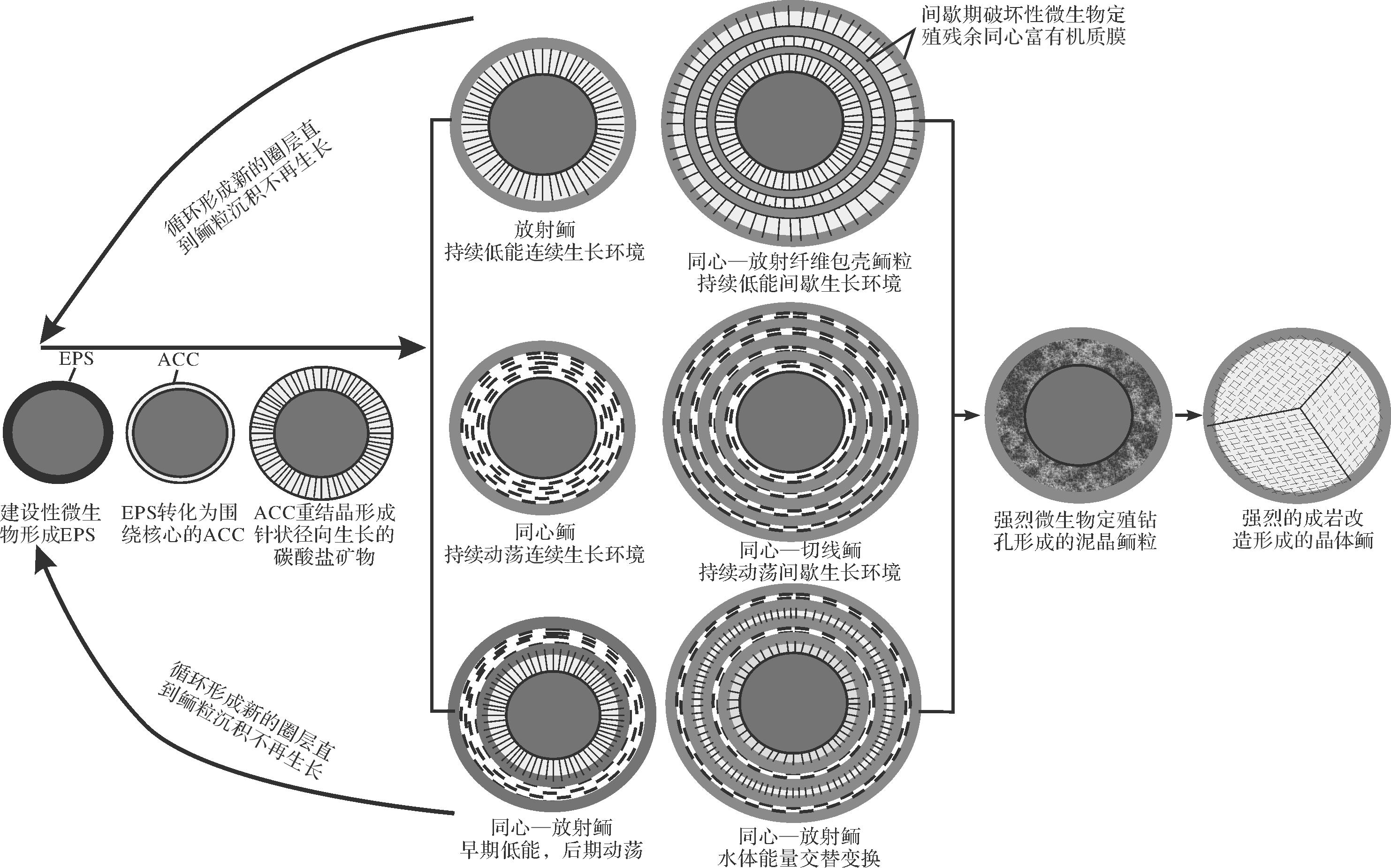
鲕粒的形成不是单一的物理化学机制或者微生物机制,微生物通过调节局部碳酸钙饱和度及碱度促进了鲕粒圈层的形成,而水动力的变化改变了鲕粒圈层结构,使得鲕粒类型丰富化,鲕粒圈层中明显的有机质圈层是鲕粒在静止阶段被微生物定殖的结果,鲕粒沉积后微生物钻孔和强烈的成岩改造可以使鲕粒圈层进一步模糊。结合前人提出的鲕粒形成模式,本文将鲕粒形成分为几个阶段(图4):1)稳定阶段围绕原始核心的光合作用微生物及硫酸盐还原细菌等建设性生物群落形成生物膜,并分泌产生细胞胞外聚合物质(EPS);2)活跃阶段鲕粒表面形成具有相对较高的Mg/Ca比的非晶质钙碳酸盐(ACC);3)稳定阶段ACC重结晶形成围绕核心径向生长的针状碳酸盐矿物,构成鲕粒圈层,该时期如果水体环境较为动荡,形成的径向生长的碳酸盐矿物被水动力改造成切线状或者杂乱分布结构,再重复第(1)和(2)步;如果水动力相对稳定且较弱,重复第(1)和(2)步形成新的径向生长的鲕粒圈层。因此,认为鲕粒径向生长的放射状圈层结构是最初始的原生圈层结构,而切线状和杂乱圈层结构是径向放射状圈层被水动力改造的结果。

图 4 不同类型鲕粒形成过程
Figure 4. Different types of ooid formation process
-
不同鲕粒在新的圈层形成前必须经过生物膜—EPS—ACC—矿化成针状碳酸盐矿物这四个过程,新圈层形成后根据水动力条件及鲕粒生长的间歇性形成几种典型的鲕粒类型(图4):1)持续低能连续生长环境,鲕粒圈层围绕核心持续径向周期性生长,生物膜后期难以保留,形成典型的放射鲕;2)持续低能间歇生长环境,鲕粒圈层围绕核心径向周期性间歇生长,在鲕粒生长间歇阶段,被大量微生物定殖,后期残余的有机质形成明显的同心状分布的富有机质层,这种情况下形成的鲕粒表现为径向生长的放射状圈层和富有机质同心圈层周期性交替,形成同心—径向放射状包壳鲕(多数学者定义的地质历史时期的同心鲕);3)持续动荡连续生长环境,ACC矿化形成围绕核心的径向生长的碳酸盐矿物被水动力剪切作用改造成围绕核心的切线状,周期性的切线状圈层包裹鲕粒核心形成典型的同心鲕;4)持续动荡间歇生长环境,切线圈层被间歇期微生物定殖形成的黑色富有机质同心圈层分割在不同的小层内,形成同心—切线包壳鲕;5)鲕粒形成早期处于静水环境,连续生长径向放射圈层,后期经过短暂间歇环境变的动荡形成连续的切线状圈层,构成典型的同心—放射鲕;6)鲕粒在形成后,受生物定殖强烈的钻孔影响,原始圈层被破坏,形成泥晶鲕;7)鲕粒沉积后期遭受强烈的成岩作用改造,尤其是重结晶作用,形成多晶鲕/单晶鲕,最外层被富含有机质的泥晶套包裹(图4)。
同心鲕和放射鲕在后期成岩作用影响下均可以表现为“花瓣状”结构,在偏光显微镜正交光下显示明显的十字消光,因此不能将十字消光的鲕粒定义为放射鲕,需要结合沉积背景及附近地层鲕粒类型综合判断。同心鲕特指以切线结构圈层为主的鲕粒,放射鲕受频繁间歇生长发育的黑色残余有机质圈层也可以表现为明显的同心圈层,本文建议将这种鲕粒定义为同心—径向放射状包壳鲕。
-
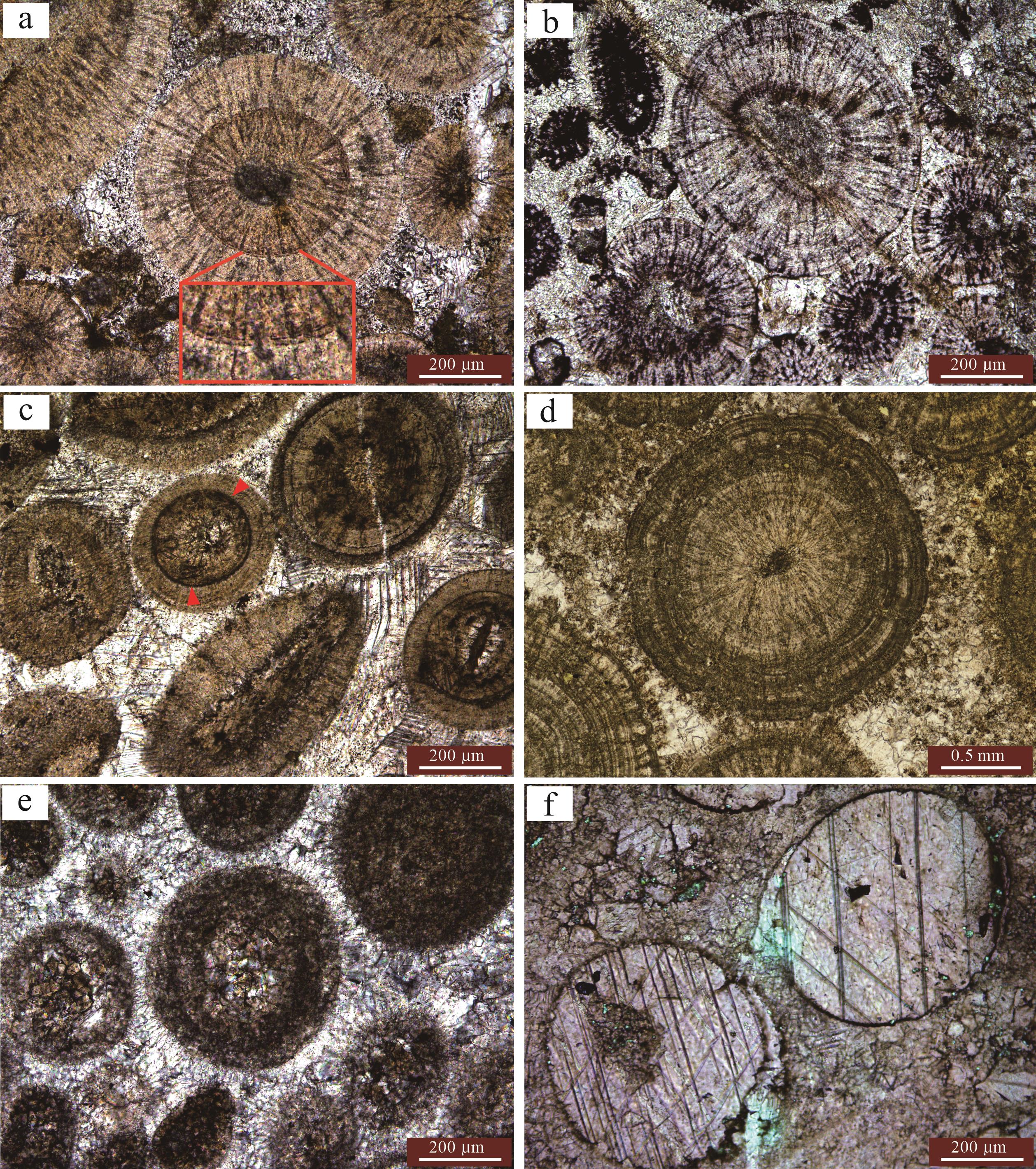
北京西山中寒武统广泛发育厚层的鲕粒石灰岩,鲕粒类型丰富多样,运用上文建立的鲕粒形成模式,可以很好地解释每一种鲕粒的形成过程。受沉积过程中水体能量频繁变化的影响,单纯的放射鲕和同心鲕是基本不发育的,普遍为放射状结构和同心状结构及富有机质膜组成的复合结构(图5a~d)。鲕粒在持续生长过程中围绕鲕粒的生物膜是很难保存的,现在所能观察到的黑色富有机质膜均说明鲕粒在生长过程中经历了长时间的埋藏间歇。鲕粒在间歇期能否被再次搅动悬浮于水体中继续生长决定了鲕粒最终的粒径。鲕粒经历了一次明显的间歇,再次搅动生长构成了最外面的圈层结构,因此该鲕粒的粒径也比周围未被搅动的鲕粒大(图5a)。鲕粒生长的间歇期可以是多次的,该鲕粒在生长后期经历了频繁的间歇生长阶段,但单次的生长规模均较小,形成了最外层的同心包壳结构(图5b)。与鲕粒二次生长环境为持续低能相比,鲕粒二次生长环境为典型的动荡高能环境,形成了明显的同心结构(图5b),但这种高能环境并不是持续稳定的,外部圈层中频繁发育的富有机质膜就是证据。泥晶鲕(图5e)、晶体鲕(图5f)为原始鲕粒受微生物、成岩作用影响形成的次生结构。

图 5 北京西山下寒武统典型鲕粒
Figure 5. Typical ooids of the Lower Cambrian in the western mountains of Beijing
-
鲕粒作为碳酸盐岩中一种重要的颗粒,关于其成因模式一直是地质学研究的重点、热点问题,鲕粒的形成也不是单一的物理化学机制或者微生物机制。本文通过对鲕粒研究综述和实例分析,认为鲕粒新圈层的形成是微生物介导的,鲕粒圈层内部碳酸盐矿物的分布形态受沉积水体能量控制,并建立了多类型鲕粒成因模式,对进一步分析鲕粒成因机制和环境指示意义具有一定的参考性。鲕粒形成过程是复杂的,分布的沉积水体环境也是多样的,如何精细地揭示微生物在鲕粒圈层形成过程中发挥的作用还需要进一步深化。
Research Progress on the Formation of Ooids
-
摘要: 鲕粒不仅是古环境的重要指示标志,由其构成的鲕粒石灰岩通常也是重要的油气储集层。但自鲕粒被发现一个多世纪以来,关于其无机成因还是有机成因就一直存在争议。分别系统归纳了典型鲕粒的物理化学成因及微生物成因、鲕粒的圈层结构类型及成因,结合前人研究成果建立了新的鲕粒成因模式,解释了常见鲕粒的形成过程。对鲕粒研究综述和实例分析认为,鲕粒新圈层的形成是微生物介导的,鲕粒圈层形态是受沉积水体能量控制的,光合微生物及硫酸盐还原细菌等建设性微生物群落均促进了鲕粒圈层的沉淀;鲕粒的圈层结构主要为径向生长的放射状、随机分布的杂乱状和切线状,在地质历史时期观察到的其他鲕粒结构均是受活跃的破坏性钻孔生物及不同程度成岩作用改造的结果;鲕粒最原始的结构是径向生长的放射状,是在低能静水环境中由非晶质钙碳酸盐(ACC)矿化形成的,随机分布结构及切线状结构是被水动力剪切改造形成的结构,鲕粒中明显的暗色同心薄层是鲕粒生长过程中间歇性出现的微生物群落建设性或破坏性定殖后生物矿化的产物,受水动力影响这三种结构既可以单独发育,也可以组合出现,构成了鲕粒形态的多样性;核心供给速率大小、沉积水体碳酸钙饱和度高低、沉积水体能量强弱及建设性微生物群落活跃程度共同控制了鲕粒粒径大小。Abstract: Oolitic limestone is a significant indicator of the paleoenvironment, and also provides an important hydrocarbon reservoir. However, ever since ooids were first recognized more than a century ago their origin, whether organic or inorganic, has been controversial. This study systematically summarizes the physicochemical and microbiological genesis of typical ooids and their structural types. A new formation mechanism for ooids is proposed, based on previous research reports. The research review and case analysis suggests that microorganisms mediate the formation of new oolitic circles. The distribution of carbonate minerals in the oolitic circles is influenced by the energy of the sedimentary water body, and constructive microbial communities (e.g., photosynthetic microorganisms and sulfate reduction bacteria) promote the precipitation of the oolitic circles. The circular structure is mainly radial, randomly distributed and tangential. Other observed oolitic structures have all been modified by the action of destructive drilling organisms and different degrees of diagenesis. The most primitive structure of an ooid is radial growth formed by the mineralization of amorphous calcium carbonate (ACC) in a low-energy, static water environment. The random distribution and tangential structure are the result of hydrodynamic shearing. The obvious dark concentric thin layers observed in ooids are the product of biomineralization of the microbial community that appears intermittently during the growth of the ooid after constructive or destructive colonization. These three structures develop either individually or in combination depending on the hydrodynamics of their environment. The core supply rate, the calcium carbonate saturation and energy of the sedimentary water body, and the action of the constructive microbial community, all influence the size of the oolitic particle size.
-
Key words:
- ooid /
- oolitic layer /
- origin process /
- oolitic size
-

-
[1] Sandberg P A. An oscillating trend in Phanerozoic non-skeletal carbonate mineralogy[J]. Nature, 1983, 305(5929): 19-22. [2] Opdyke B N, Wilkinson B H. Carbonate mineral saturation state and cratonic limestone accumulation[J]. American Journal of Science, 1993, 293(3): 217-234. [3] Wilkinson B H, Owen R M, Carroll A R. Submarine hydrothermal weathering, global eustasy, and carbonate polymorphism in Phanerozoic marine oolites[J]. Journal of Sedimentary Research, 1985, 55(2): 171-183. [4] 郭芪恒,金振奎,安益辰,等. 北京下苇甸地区张夏组沉积环境及沉积模式[J]. 沉积学报,2019,37(1):40-50. Guo Qiheng, Jin Zhenkui, An Yichen, et al. Study on sedimentary environment and patterns of the Cambrian Zhangxia Formation at Xiaweidian, Beijing[J]. Acta Sedimentologica Sinica, 2019, 37(1): 40-50. [5] 郭芪恒,金振奎,史书婷,等. 鲕粒粒度特征及其指示意义:以北京西山下苇甸寒武系张夏组剖面为例[J]. 沉积学报,2020,38(4):737-746. Guo Qiheng, Jin Zhenkui, Shi Shuting, et al. Characteristics of ooid size and its environmental significance: A case study from the Cambrian Zhangxia Formation at Xiaweidian outcrop, Beijing[J]. Acta Sedimentologica Sinica, 2020, 38(4): 737-746. [6] 李飞,武思琴,刘柯. 鲕粒原生矿物识别及对海水化学成分变化的指示意义[J]. 沉积学报,2015,33(3):500-511. Li Fei, Wu Siqin, Liu Ke. Identification of ooid primary mineralogy: A clue for understanding the variation in paleo-oceanic chemistry[J]. Acta Sedimentologica Sinica, 2015, 33(3): 500-511. [7] 李开开,张学丰,贺训云,等. 川东北飞仙关组白云岩化作用对鲕粒滩储层的孔隙改造效应[J]. 石油与天然气地质,2018,39(4):706-718. Li Kaikai, Zhang Xuefeng, He Xunyun, et al. Modification of dolomitization on pores in oolitic shoal reservoirs of the Feixianguan Formation in the northeastern Sichuan Basin[J]. Oil & Gas Geology, 2018, 39(4): 706-718. [8] 刘冉,霍飞,王鑫,等. 普光气田下三叠统飞仙关组碳酸盐岩储层特征及主控因素分析[J]. 中国石油勘探,2017,22(6):34-46. Liu Ran, Huo Fei, Wang Xin, et al. Characteristics and main controlling factors of Lower Triassic Feixianguan Formation carbonate reservoir in Puguang gas field[J]. China Petroleum Exploration, 2017, 22(6): 34-46. [9] 李宏涛. 台内鲕粒滩气藏成藏过程与模式:以川东北河坝地区下三叠统飞仙关组三段为例[J]. 石油勘探与开发,2016,43(5):723-732. Li Hongtao. Accumulation process and pattern of oolitic shoal gas pools in the platform: A case from member 3 of Lower Triassic Feixianguan Formation in the Heba area, northeastern Sichuan Basin[J]. Petroleum Exploration and Development, 2016, 43(5): 723-732. [10] 梅冥相, Latif K,孟晓庆,等. 鲕粒滩中光合作用生物膜构建的高能核形石:以辽西葫芦岛三道沟剖面寒武系张夏组为例[J]. 地质学报,2020,94(4):999-1016. Mei Mingxiang, Latif K, Meng Xiaoqing, et al. High-energy oncoids within the ooid-grained bank built by photosynthetic biofilms: A case study of the Cambrian Zhangxia Formation at the Sandaogou section of Huludao city in the western part of Liaoning province[J]. Acta Geologica Sinica, 2020, 94(4): 999-1016. [11] 杨仁超,樊爱萍,韩作振,等. 核形石研究现状与展望[J]. 地球科学进展,2011,26(5):465-474. Yang Renchao, Fan Aiping, Han Zuozhen, et al. Status and prospect of studies on oncoid[J]. Advances in Earth Science, 2011, 26(5): 465-474. [12] Sorby H C. The structure and origin of limestones[J]. Proceedings of the Geological Society of London, 1879, 35: 56-95. [13] Rankey E C, Reeder S L. Holocene ooids of Aitutaki atoll, cook islands, South Pacific[J]. Geology, 2009, 37(11): 971-974. [14] Pacton M, Ariztegui D, Wacey D, et al. Going nano: A new step toward understanding the processes governing freshwater ooid formation[J]. Geology, 2012, 40(6): 547-550. [15] Plee K, Ariztegui D, Martini R, et al. Unravelling the microbial role in ooid formation-results of an in situ experiment in modern freshwater Lake Geneva in Switzerland[J]. Geobiology, 2008, 6(4): 341-350. [16] Davies P J, Bubela B, Ferguson J. The formation of ooids[J]. Sedimentology, 1978, 25(5): 703-730. [17] Royal Society. Ocean acidification due to increasing atmospheric carbon dioxide: London. The Royal Society, Policy Document, 2005, 12/05, 57 p. [18] Duguid S M A, Kyser T K, James N P, et al. Microbes and ooids[J]. Journal of Sedimentary Research, 2010, 80(3/4): 236-251. [19] Trower E J, Cantine M D, Gomes M L, et al. Active ooid growth driven by sediment transport in a high-energy shoal, little ambergris cay, Turks and Caicos islands[J]. Journal of Sedimentary Research, 2018, 88(9): 1132-1151. [20] Folk R L, Lynch F L. Organic matter, putative nannobacteria and the formation of ooids and hardgrounds[J]. Sedimentology, 2001, 48(2): 215-229. [21] Summons R E, Bird L R, Gillespie A L, et al. Lipid biomarkers in ooids from different locations and ages: Evidence for a common bacterial flora[J]. Geobiology, 2013, 11(5): 420-436. [22] Tang D J, Shi X Y, Shi Q, et al. Organomineralization in Mesoproterozoic giant ooids[J]. Journal of Asian Earth Sciences, 2015, 107: 195-211. [23] Tan Q, Shi Z J, Tian Y M, et al. Origin of ooids in ooidal-muddy laminites: A case study of the Lower Cambrian Qingxudong Formation in the Sichuan Basin, South China[J]. Geological Journal, 2017, 53(5): 1716-1727. [24] Diaz M R, Swart P K, Eberli G P, et al. Geochemical evidence of microbial activity within ooids[J]. Sedimentology, 2015, 62(7): 2090-2112. [25] Diaz M R, Eberli G P. Decoding the mechanism of formation in marine ooids: A review[J]. Earth-Science Reviews, 2019, 190: 536-556. [26] Diaz M R, Eberli G P, Blackwelder P, et al. Microbially mediated organomineralization in the formation of ooids[J]. Geology, 2017, 45(9): 771-774. [27] 梅冥相. 鲕粒成因研究的新进展[J]. 沉积学报,2012,30(1):20-32. Mei Mingxiang. Brief introduction on new advances on the origin of ooids[J]. Acta Sedimentologica Sinica, 2012, 30(1): 20-32. [28] Siahi M, Hofmann A, Master S, et al. Carbonate ooids of the mesoarchaean pongola supergroup, South Africa[J]. Geobiology, 2017, 15(6): 750-766. [29] Brehm U, Krumbein W E, Palinska K A. Biomicrospheres generate ooids in the laboratory[J]. Geomicrobiology Journal, 2006, 23(7): 545-550. [30] Brehm U, Palinska K A, Krumbein W E. Laboratory cultures of calcifying biomicrospheres generate ooids: A contribution to the origin of oolites[J]. Carnets De Géologie, 2004, CG2004(L03): 1-6. [31] 宋文天,刘建波. 碳酸盐鲕粒包壳结构研究综述[J]. 古地理学报,2020,22(1):147-160. Song Wentian, Liu Jianbo. A review of cortical structures of carbonate ooids[J]. Journal of Palaeogeography, 2020, 22(1): 147-160. [32] Rao V P, Milliman J D. Relict ooids off northwestern India: Inferences on their genesis and Late Quaternary sea level[J]. Sedimentary Geology, 2017, 358: 44-50. [33] Sandberg P A. New interpretations of Great Salt Lake ooids and of ancient non‐skeletal carbonate mineralogy[J]. Sedimentology, 1975, 22(4): 497-537. [34] Simone L. Ooids: A review[J]. Earth-Science Reviews, 1980, 16: 319-355. [35] 周瑶琪,张晗,张振凯. 海相碳酸盐鲕粒形成过程的模拟实验研究[J]. 中国石油大学学报(自然科学版),2017,41(3):23-30. Zhou Yaoqi, Zhang Han, Zhang Zhenkai. Experiment study of synthesis for marine carbonate ooids genesis[J]. Journal of China University of Petroleum (Edition of Natural Science), 2017, 41(3): 23-30. [36] Sumner D Y, Grotzinger J P. Numerical modeling of ooid size and the problem of Neoproterozoic giant ooids[J]. Journal of Sedimentary Petrology, 1993, 63(5): 974-982. [37] 段雄,时志强,金鑫,等. 巨鲕的微生物成因:来自重庆石柱地区下寒武统的证据[J]. 古地理学报,2015,17(2):241-248. Duan Xiong, Shi Zhiqiang, Jin Xin, et al. Microbial cause for giant ooids: Evidence from the Lower Cambrian in Shizhu area, Chongqing[J]. Journal of Palaeogeography, 2015, 17(2): 241-248. [38] Trower E J, Lamb M P, Fischer W W. Experimental evidence that ooid size reflects a dynamic equilibrium between rapid precipitation and abrasion rates[J]. Earth and Planetary Science Letters, 2017, 468: 112-118. [39] Li F, Gong Q L, Burne R V, et al. Ooid factories operating under hothouse conditions in the earliest Triassic of South China[J]. Global and Planetary Change, 2019, 172: 336-354. -

 点击查看大图
点击查看大图
图(5)
计量
- 文章访问数: 1856
- HTML全文浏览量: 454
- PDF下载量: 480
- 被引次数: 0

