


-
白云岩(石)的成因一直是沉积地质学领域的一个热点和难点问题,尽管对它的研究已经超过200年,但白云岩的成因问题仍然是困扰地质学家的谜团[1]。首先,白云岩在地质历史时期的碳酸盐岩台地中分布广泛,但在对白云石过饱和的现代海洋中却罕见[2⁃3];其次,在无机实验室条件下难以沉淀出有序白云石。因此,人们难以利用现代实例或合成实验解释地质历史时期广泛分布的白云岩的成因问题。在Ca2+-Mg2+-CO2-H2O平衡体系中,白云石对现代正常海水(平均盐度和Mg2+/Ca2+比值)在热力学上是稳定的,然而现代海水中却并没有产生广泛的白云石沉淀,这一理论计算和实际观测的矛盾被归因于白云石的形成受到化学动力学因素的限制[4⁃9]。动力学因素可能从以下几方面影响白云石的形成[10]:(1)Mg2+的水合作用强于Ca2+,更容易形成富钙碳酸盐矿物,而抑制白云石的形成[7];(2)较低的CO
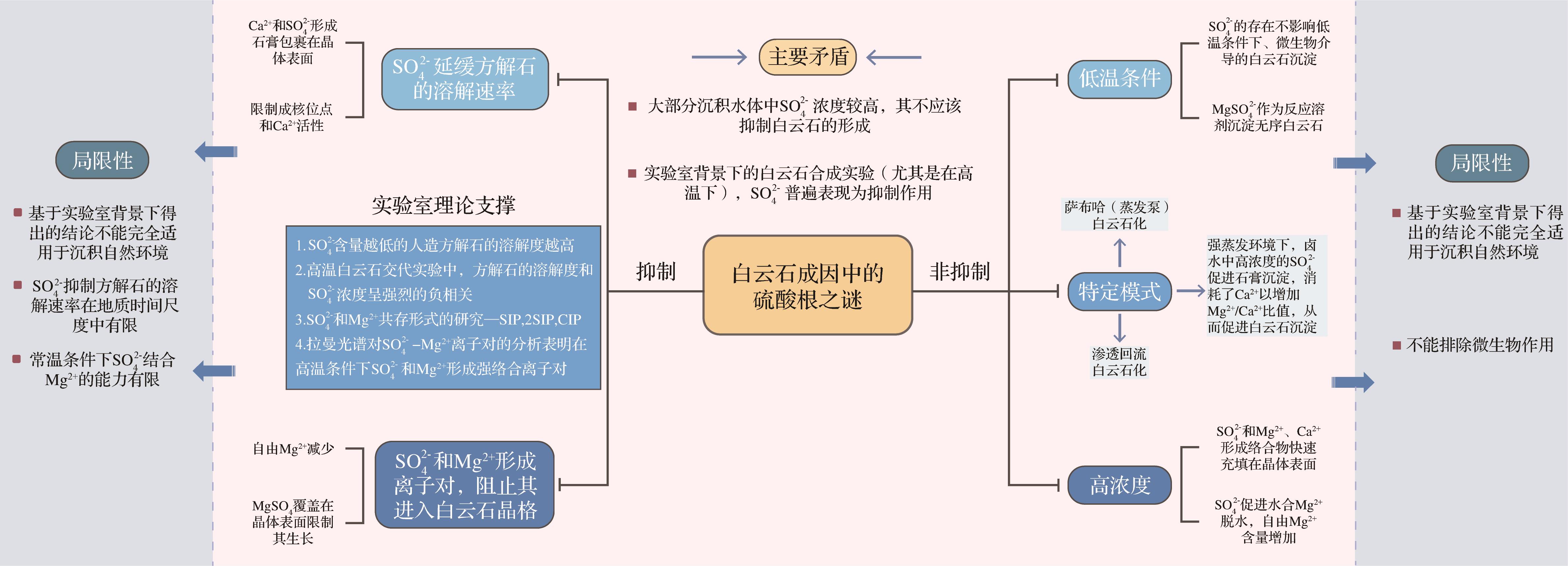
在“白云岩问题”研究中,SO
对“硫酸根之谜”的探索与揭示,不仅是解决“白云岩问题”的关键一环,而且在一定程度上影响人们对古海洋环境演化的认识。白云岩作为地质记录中常见的碳酸盐岩,其丰度变化可能与古海洋环境演化有着密切联系[40⁃44]。长期以来,硫酸根抑制白云石形成的观点被普遍接受,并被用于解释元古代广泛发育的白云岩与低海水硫酸根离子浓度之间的负相关关系[42]。近年来,地质历史时期白云石的形成与海水性质(尤其是海水硫酸根浓度)、极端地质事件之间的关系引起了地质学家的关注,如Li et al.[43]提出在二叠纪—三叠纪之交,全球海洋在化学跃变层附近发生了一次广泛的白云岩沉积事件,对应高温、缺氧及硫化的极端海洋环境,并伴随着一次快速而显著的海水硫酸盐含量降低,因此,将此次白云岩事件与缺氧海水环境下硫酸盐还原菌的爆发联系起来;此外,Li et al.[44]对显生宙白云石含量进行了统计分析,并结合古生物数据及硫同位素数据,发现白云石含量的峰值与硫同位素快速上升时期相对应,硫同位素的快速上升反映了微生物硫酸盐还原速率的增加,进而指示了海洋缺氧及硫化事件的发生。这一发现间接证明了海水中的SO
“硫酸根之谜”作为“白云岩问题”中未解决的争议之一,前人尚未对其具体的影响机理和研究现状进行系统归纳和总结。因此,本文以白云石形成过程中的“硫酸根之谜”为切入点,对近六十年来关于SO
-
人们对白云石成因中“硫酸根之谜”的研究最早可追溯到半个多世纪前,Liebermann[34]利用Frear et al.[47]的研究作为理论前提,提出溶解的硫酸钙通过降低方解石的溶解度进而抑制白云石的形成。然而,Liebermann[34]未将这种抑制作用锁定在SO
在SO

图 1 白云石成因中“硫酸根之谜”的研究历程及主流观点
Figure 1. Research history and mainstream views of the "sulfate enigma" of dolomite genesis
然而,SO
近二十年来,微生物因素的加入为“硫酸根之谜”的研究注入了新的活力。1997年,Vasconcelos et al.[15]提出了首个被学界认可的微生物诱(介)导白云石沉淀的模型。该模型提出初期,被认为是SO
-
Baker et al.[4]首次提出硫酸根是白云石形成过程中的抑制剂,他们认为即使很低浓度的SO

图 2 白云石形成过程中硫酸根作用的不同观点
Figure 2. Impacts of sulfate on the formation of dolomite from diverse perspectives
-
Liebermann[34]最初认为溶解的CaSO4主要通过降低方解石的溶解度来抑制白云石的形成,但他没有利用进一步的实验来佐证这种观点,也没有将这种抑制作用锁定在SO
表 1 以SO
Table 1. Dolomite synthesis experiments with SO
实验类型 实验温度 分析方法 关键实验数据 实验结论 参考文献 白云石低温合成实验 40 ℃~50 ℃ X射线衍射 实验编号 是否含CaSO4/SO 镁方解石或白云石的百分重量 沉淀物质类型 CaSO4抑制白云石的形成 Liebermann[34] 29、30、31 是 无 无 24 否 很少(little) 镁方解石 52 是 微量(traces) 方解石—白云石中间体 53 否 5% 无序白云石 55 否 5% 无序白云石 57 否 微量(traces) 有序白云石 高温热液实验 200 ℃ X射线衍射扫描电镜 初始SO 沉淀物质类型 SO Baker et al.[4] 0 白云石 1 原白云石(含43 mol % Mg)>>方解石 2 方解石>>原白云石(含45 mol % Mg) 3 方解石>>原白云石(含46 mol % Mg) 4 方解石 白云石高温交代实验 215 ℃ X射线衍射 初始SO 沉淀物质类型 白云石含量最高对应的交代时间 SO Morrow et al.[36] 0 白云石(74%)+水镁石(26%) 7天 4 水镁石(53%)+白云石(28%)+硬石膏(19%) 14天 20 无白云石出现 — 微生物白云石低温诱导实验采用不消耗SO <50 ℃ X射线衍射扫描电镜 温度 SO 沉淀的白云石含量单位:SI=log(IAP/Ksp) 温度 SO 沉淀的白云石含量单位:SI=log(IAP/Ksp) SO Sánchez-Román et al.[16] 25 ℃ 0 -11.10 35 ℃ 0 -11.98 14 -13.46 14 -14.18 28 -15.72 23 -16.34 56 -17.07 56 -17.49 硫化物催化白云石交代实验 室温 X射线衍射扫描电镜 硫化物浓度/mM 沉淀物质类型及含量 硫化物(HS-、H2S)可以催化白云石的形成/SO Zhang et al.[23] 4.0 霞石(14%)+方解石粉末(14%)+高镁方解石(72%) 6.0 霞石(18%)+方解石粉末(26%)+高镁方解石(56%) 7.2 霞石(6%)+方解石粉末(25%)+无序白云石(69%) 8.4 霞石(5%)+方解石粉末(22%)+无序白云石(73%) -
Slaughter et al.[37]研究认为,溶液中的SO
Mg2++SO
水合离子→双溶剂分离离子对→溶剂共用离子对→强络合的接触离子对 (1) 此后Akilan et al.[53]的研究表明,随着温度的升高,2SIP和SIP的稳定性会急速减弱,在200 ℃的高温下,2SIP和SIP几乎不存在。Wang et al.[49]用拉曼光谱对MgCl2-NaCl-MgSO4-H2O体系中的Mg2+-SO
从上述研究中可以看出,SO
此外,还需要考虑不同温度下SO
-
尽管有大量研究认为SO
表 2 现代白云石沉淀的部分沉积环境及相应水体中的SO
Table 2. Sedimentary environments where dolomite is still precipitated today and the concentrations of sulfate in corresponding water bodies
序号 沉积环境 位置 时代 SO 参考文献 1 Deep Springs湖 美国西部 现代、全新世 211.4~422.8 Jones[59] 2 Great Salt湖 美国西部 现代、全新世、更新世 92~130 Jagniecki et al.[60] 3 Devils湖 美国中北部 现代、全新世 11.97 Engstrom et al. [61] 4 Waldsea湖 加拿大西部 现代、全新世 0.130~0.313 Last et al.[62] 5 Freefight湖 加拿大西部 现代、全新世 666~957 Lyons et al.[63] 6 British Columbia湖 加拿大西部 现代、全新世 0.949 Strang et al.[64] 7 Coorong湖 澳大利亚南部 现代、全新世 100.7~589.5 Wright et al.[65] 8 CurtinSpring湖 澳大利亚中部 更新世 326.2~499.7 Jacobson et al.[66] 9 Frome湖 澳大利亚中部 全新世 13.52 Pirlo et al.[67] 10 Eyre湖 澳大利亚中部 更新世 9.55~137.42 Tweed et al.[68] 11 Macleod湖 澳大利亚西部 现代、全新世 >0.208 Papineau et al.[69] 12 Balaton湖 匈牙利西部 全新世 0.616~1.367 Simon et al.[70] 13 Spanish playas湖 西班牙中部和东北部 现代、全新世 932.774 Lopez et al.[71] 14 Baza Basin湖 西班牙东南部 更新世 14.21~27.49 Hidalgo et al.[72] 15 Tuz湖 土耳其中部 全新世、更新世 63.49~705.5 Camur et al.[73] 16 Turkish playas湖 土耳其中部 现代 168.5 Kuşcu et al.[74] 17 Van湖 土耳其东部 全新世 17.6~28.9 Reimer et al.[75] 18 Urmia湖 伊朗西北部 全新世 147.5 Alipour[76] 19 Balkhash湖 哈萨克斯坦东部 现代 23.4 Dzhetimov et al.[77] 20 Doroninskoye湖 俄罗斯东西伯利亚 现代 1.845 Borzenko et al.[78] 21 吉布胡郎图湖 中国内蒙古 全新世 117.5 Liu et al.[79] 22 察尔汗盐湖 柴达木盆地 全新世 0.58~5.8 Fan et al.[80] 23 Bogoria湖 非洲东部 更新世 0.11~1.5 Cioni et al.[81] 24 Turkana湖 非洲东部 更新世 0.4 Yan et al.[82] 25 Kivu湖 非洲卢旺达西北部 全新世 <0.1 Hategekimana et al.[83] 26 Sayram湖 亚洲中部 早全新世 16.88 Cheng et al.[84] 27 Lagoa Vermelha湖 巴西南大河洲 现代 41~60 Warthmann et al.[85] 28 Lagoa Vermelha湖 巴西南大河洲 现代 50 van Lith et al.[86] 29 Brejo do Espinho湖 巴西里约热内卢州 现代 69 van Lith et al.[86] 30 Seawater — — ≈28.125 Corzo et al.[87] 29 Brennan et al.[88] -
早在1961年,Siegel[92]利用MgSO4作为反应试剂之一,在低温下(25 ℃)沉淀出了无序白云石。之后Rivadeneyra et al.[56⁃58]使用浓度为15%和20%的人工海水(SO
-
早在1962年,Wells[94]报道了波斯湾地区蒸发浓缩形成的与石膏共生的白云岩。而后Chilingar et al.[95]研究证明了白云石可以在饱和的氯化物—硫酸盐混合溶液中沉淀,为蒸发泵白云石的形成提供了先导理论支撑。1969年,Hsü et al.[96]正式称其为蒸发泵(萨布哈)作用,其原理为疏松沉积物中的孔隙水通过强烈蒸发进行浓缩,而海水通过毛细管作用补给到疏松多孔沉积物中,久而久之,海水变成高SO
-
Brady et al.[39]的研究认为低浓度(小于5 mM)的SO
为了验证Brady et al.[39]的结论在自然界沉积水体中的适用性,本文对现代白云石发育的部分沉积环境及相应水体中的SO
综上所述,在特定的条件下,SO
-
随着微生物白云石模式的提出,多个学者发现一些细菌对SO
目前的研究表明,硫酸盐还原菌(SRB)、甲烷生成菌、甲烷氧化菌(MOB)和嗜盐好氧细菌是介导白云石形成的4种主要微生物[18,103]。其中SRB和MOB的代谢活动与SO
-
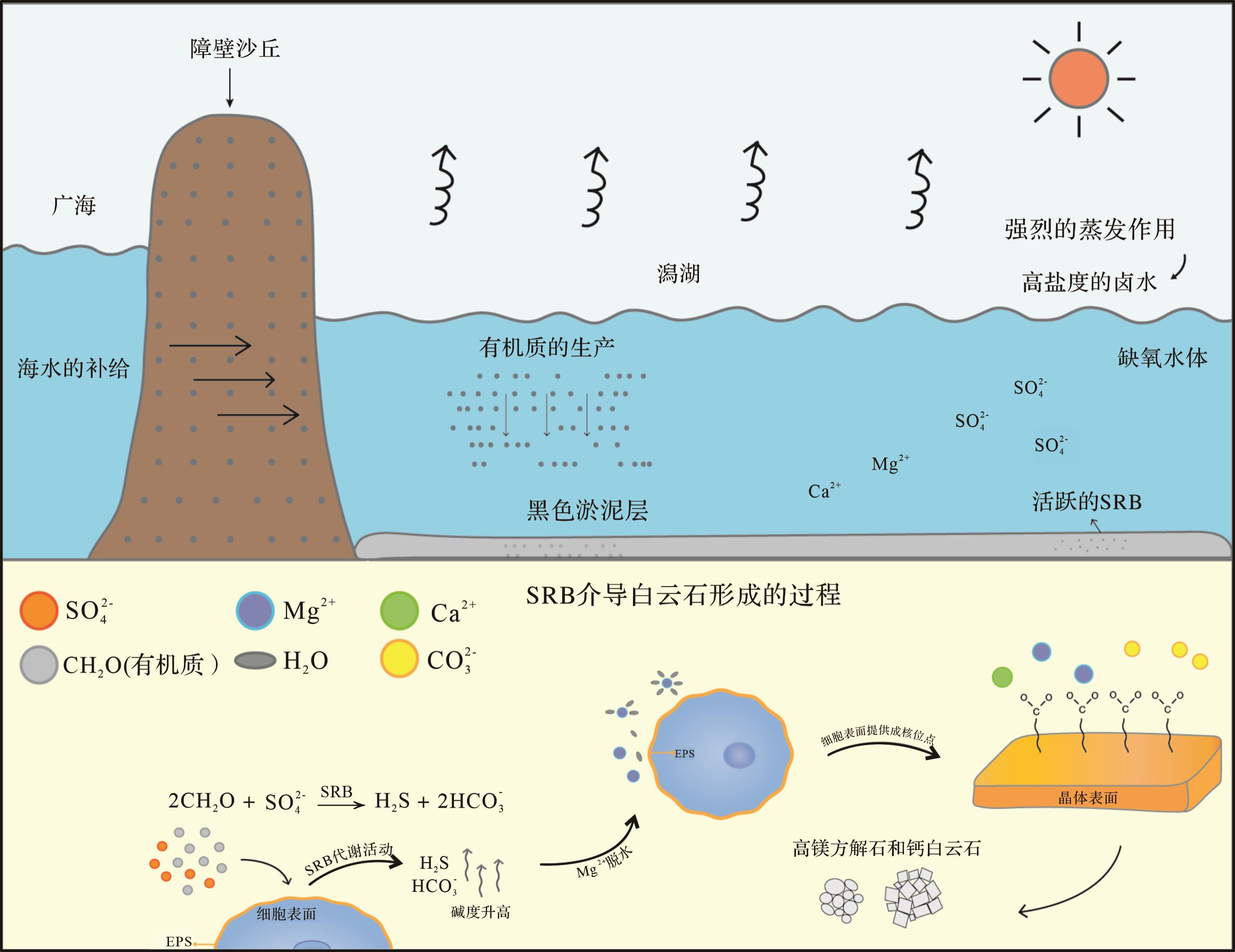
Vasconcelos et al.[15]在巴西东海岸的Lagoa Vermelha潟湖缺氧还原性黑色富有机质沉积物中发现了现代白云石,并利用采自该潟湖的硫酸盐还原菌Desulforvibrio在实验室厌氧条件下沉淀出了低有序度的白云石,从而正式提出了硫酸盐还原菌介导的微生物白云石模式。如图3所示,在旱季,强烈的蒸发作用使得湖中水位降低,海水由障壁沙丘流入湖中进行补充,湖底缺氧黑色淤泥层中(淤泥主要是碳酸盐泥、细菌、有机质和水的混合物)的SRB非常活跃,SRB的代谢活动创造了利于白云石形成的过饱和条件(图3),即细胞周围聚集了大量的Mg2+、Ca2+、HCO

如今,有SO
-
大量研究表明,BSR和AOM通过降低SO
部分学者认为,SRB和MOB促进白云石沉淀的本质是因为移除了SO
此外,Zhang et al.[23]提出硫化物可以催化白云石的形成(表1),他们的研究表明溶解的H2S和HS-中的H会和方解石中CO
从另一种角度来看,SO
BSR:2CH2O(有机质)+SO (2) AOM:CH4+SO (3) 所以从动力学角度而言,必须连续供应SO
综上,在微生物白云石模式中SO
另外,Deng et al.[93]研究提出SO
-
类白云石矿物是指具有与白云石(CaMg(CO3)2)阳离子有序结构类似的双金属矿物[116]。目前已知的类白云石碳酸盐矿物主要有锰白云石、铁白云石、锌白云石、钡白云石、铅白云石、镉白云石和锶白云石这七种(表3)。由于白云石低温合成实验的失败很大程度上限制了人们对白云石成因的认识[5,34],当“白云岩问题”始终无法被正面突破时,对类白云石矿物的研究便是我们从侧面解决“白云岩问题”的途径之一。钡白云石和铅白云石作为两种具有代表性的类白云石碳酸盐矿物,不仅可以在低温下无机合成,而且结构中含有与白云石等量的Mg2+。因此,研究SO
表 3 主要类白云石碳酸盐矿物及SO
Table 3. Major carbonate mineral analogues of dolomite and the role of sulfate in their formation
矿物名称 化学式 空间群 能否无机合成 SO 参考文献 锰白云石(Kutnohorite) CaMn(CO3)2 R-3 难以合成有序锰白云石 — Farkas et al.[117] 铁白云石(Ankerite) CaFe(CO3)2 R-3 未能成功合成 — Beran et al. [118] 锌白云石(Minrecordite) CaZn(CO3)2 R-3 未能成功合成 — Rosenberg et al. [119] 钡白云石(Norsethite) BaMg(CO3)2 R32 室温下即可成功合成 抑制 Lippmann[120] R-3m Effenberger et al. [121]; Secco et al. [122] R-3c Effenberger et al.[123] 铅白云石 PbMg(CO3)2 R32 室温下即可成功合成 不抑制 Lippmann[6,124] 镉白云石 CdMg(CO3)2 R-3 — — Goldsmith[125] 锶白云石 SrMg(CO3)2 R32 高温高压下可合成 不抑制 Froese[126] 钡白云石和铅白云石与钙白云石几乎存在一样的晶体结构:在c轴上都表现为阳离子交替排序的“超结构”分层[35,127]。在相同的条件下,如果SO
然而,Morrow et al.[35⁃36]的理论与白云石结晶的实际情况相悖,因为CaMg(CO3)2的溶解度低于CaSO4[129],如果溶解度小的矿物先析出,CaCO3释放的Ca2+就应该先参与形成CaMg(CO3)2而不是CaSO4,那么SO
-
在“白云岩问题”研究中,SO
“硫酸根之谜”作为“白云岩问题”中仍未解决的难题之一,在未来的研究中,需重点关注以下几方面内容:(1)鉴于SO
‘Sulfate Enigma’ in Dolomite Genesis: Review and Perspectives
-
摘要: 意义 “白云岩问题”是沉积学领域最具争议的热点、难点问题之一。大量研究表明,无论是在实验室条件还是现代自然沉积环境中,都难以在低温、无机条件下直接沉淀有序白云石。前人研究认为白云石的形成是一个受动力学控制的过程,并提出了几种关键制约因素,包括镁离子的水合作用、抑制剂硫酸根的存在、成核位点及阳离子的有序化等。硫酸根在白云石形成过程中的作用一直备受关注,但存在较大争议。长期以来,硫酸根抑制白云石形成的假说被地质学家们广泛接受,并被用于解释地质历史时期白云岩丰度与海水性质的演化。但随着研究的深入,尤其是对微生物白云石成因机理的不断探索,人们对白云石形成过程中硫酸根作用的认识逐渐出现了争议,一些学者认为硫酸根并不会抑制白云石的形成。对硫酸根的这种争议性认识主要是由于:(1)现代白云石形成的自然沉积环境中SO
-
Key words:
- dolomite problem /
- kinetic barrier /
- sulfate /
- inhibiting mechanism /
- facilitating mechanism /
- microbial dolomite
-
图 1 白云石成因中“硫酸根之谜”的研究历程及主流观点
Figure 1. Research history and mainstream views of the "sulfate enigma" of dolomite genesis
图 2 白云石形成过程中硫酸根作用的不同观点
Figure 2. Impacts of sulfate on the formation of dolomite from diverse perspectives
表 1 以SO
Table 1. Dolomite synthesis experiments with SO
实验类型 实验温度 分析方法 关键实验数据 实验结论 参考文献 白云石低温合成实验 40 ℃~50 ℃ X射线衍射 实验编号 是否含CaSO4/SO 镁方解石或白云石的百分重量 沉淀物质类型 CaSO4抑制白云石的形成 Liebermann[34] 29、30、31 是 无 无 24 否 很少(little) 镁方解石 52 是 微量(traces) 方解石—白云石中间体 53 否 5% 无序白云石 55 否 5% 无序白云石 57 否 微量(traces) 有序白云石 高温热液实验 200 ℃ X射线衍射扫描电镜 初始SO 沉淀物质类型 SO Baker et al.[4] 0 白云石 1 原白云石(含43 mol % Mg)>>方解石 2 方解石>>原白云石(含45 mol % Mg) 3 方解石>>原白云石(含46 mol % Mg) 4 方解石 白云石高温交代实验 215 ℃ X射线衍射 初始SO 沉淀物质类型 白云石含量最高对应的交代时间 SO Morrow et al.[36] 0 白云石(74%)+水镁石(26%) 7天 4 水镁石(53%)+白云石(28%)+硬石膏(19%) 14天 20 无白云石出现 — 微生物白云石低温诱导实验采用不消耗SO <50 ℃ X射线衍射扫描电镜 温度 SO 沉淀的白云石含量单位:SI=log(IAP/Ksp) 温度 SO 沉淀的白云石含量单位:SI=log(IAP/Ksp) SO Sánchez-Román et al.[16] 25 ℃ 0 -11.10 35 ℃ 0 -11.98 14 -13.46 14 -14.18 28 -15.72 23 -16.34 56 -17.07 56 -17.49 硫化物催化白云石交代实验 室温 X射线衍射扫描电镜 硫化物浓度/mM 沉淀物质类型及含量 硫化物(HS-、H2S)可以催化白云石的形成/SO Zhang et al.[23] 4.0 霞石(14%)+方解石粉末(14%)+高镁方解石(72%) 6.0 霞石(18%)+方解石粉末(26%)+高镁方解石(56%) 7.2 霞石(6%)+方解石粉末(25%)+无序白云石(69%) 8.4 霞石(5%)+方解石粉末(22%)+无序白云石(73%)  下载: 导出CSV
下载: 导出CSV
表 2 现代白云石沉淀的部分沉积环境及相应水体中的SO
Table 2. Sedimentary environments where dolomite is still precipitated today and the concentrations of sulfate in corresponding water bodies
序号 沉积环境 位置 时代 SO 参考文献 1 Deep Springs湖 美国西部 现代、全新世 211.4~422.8 Jones[59] 2 Great Salt湖 美国西部 现代、全新世、更新世 92~130 Jagniecki et al.[60] 3 Devils湖 美国中北部 现代、全新世 11.97 Engstrom et al. [61] 4 Waldsea湖 加拿大西部 现代、全新世 0.130~0.313 Last et al.[62] 5 Freefight湖 加拿大西部 现代、全新世 666~957 Lyons et al.[63] 6 British Columbia湖 加拿大西部 现代、全新世 0.949 Strang et al.[64] 7 Coorong湖 澳大利亚南部 现代、全新世 100.7~589.5 Wright et al.[65] 8 CurtinSpring湖 澳大利亚中部 更新世 326.2~499.7 Jacobson et al.[66] 9 Frome湖 澳大利亚中部 全新世 13.52 Pirlo et al.[67] 10 Eyre湖 澳大利亚中部 更新世 9.55~137.42 Tweed et al.[68] 11 Macleod湖 澳大利亚西部 现代、全新世 >0.208 Papineau et al.[69] 12 Balaton湖 匈牙利西部 全新世 0.616~1.367 Simon et al.[70] 13 Spanish playas湖 西班牙中部和东北部 现代、全新世 932.774 Lopez et al.[71] 14 Baza Basin湖 西班牙东南部 更新世 14.21~27.49 Hidalgo et al.[72] 15 Tuz湖 土耳其中部 全新世、更新世 63.49~705.5 Camur et al.[73] 16 Turkish playas湖 土耳其中部 现代 168.5 Kuşcu et al.[74] 17 Van湖 土耳其东部 全新世 17.6~28.9 Reimer et al.[75] 18 Urmia湖 伊朗西北部 全新世 147.5 Alipour[76] 19 Balkhash湖 哈萨克斯坦东部 现代 23.4 Dzhetimov et al.[77] 20 Doroninskoye湖 俄罗斯东西伯利亚 现代 1.845 Borzenko et al.[78] 21 吉布胡郎图湖 中国内蒙古 全新世 117.5 Liu et al.[79] 22 察尔汗盐湖 柴达木盆地 全新世 0.58~5.8 Fan et al.[80] 23 Bogoria湖 非洲东部 更新世 0.11~1.5 Cioni et al.[81] 24 Turkana湖 非洲东部 更新世 0.4 Yan et al.[82] 25 Kivu湖 非洲卢旺达西北部 全新世 <0.1 Hategekimana et al.[83] 26 Sayram湖 亚洲中部 早全新世 16.88 Cheng et al.[84] 27 Lagoa Vermelha湖 巴西南大河洲 现代 41~60 Warthmann et al.[85] 28 Lagoa Vermelha湖 巴西南大河洲 现代 50 van Lith et al.[86] 29 Brejo do Espinho湖 巴西里约热内卢州 现代 69 van Lith et al.[86] 30 Seawater — — ≈28.125 Corzo et al.[87] 29 Brennan et al.[88]
下载: 导出CSV
表 3 主要类白云石碳酸盐矿物及SO
Table 3. Major carbonate mineral analogues of dolomite and the role of sulfate in their formation
矿物名称 化学式 空间群 能否无机合成 SO 参考文献 锰白云石(Kutnohorite) CaMn(CO3)2 R-3 难以合成有序锰白云石 — Farkas et al.[117] 铁白云石(Ankerite) CaFe(CO3)2 R-3 未能成功合成 — Beran et al. [118] 锌白云石(Minrecordite) CaZn(CO3)2 R-3 未能成功合成 — Rosenberg et al. [119] 钡白云石(Norsethite) BaMg(CO3)2 R32 室温下即可成功合成 抑制 Lippmann[120] R-3m Effenberger et al. [121]; Secco et al. [122] R-3c Effenberger et al.[123] 铅白云石 PbMg(CO3)2 R32 室温下即可成功合成 不抑制 Lippmann[6,124] 镉白云石 CdMg(CO3)2 R-3 — — Goldsmith[125] 锶白云石 SrMg(CO3)2 R32 高温高压下可合成 不抑制 Froese[126]
下载: 导出CSV
-
[1] Machel H G. Concepts and models of dolomitization: A critical reappraisal[J]. Geological Society, London, Special Publications, 2004, 235(1): 7-63. [2] Cai W K, Liu J H, Zhou C H, et al. Structure, genesis and resources efficiency of dolomite: New insights and remaining enigmas[J]. Chemical Geology, 2021, 573: 120191. [3] Fairbridge R W. The dolomite question[M]//le Blanc R J, Breeding J G. Regional aspects of carbonate deposition. New York: SEPM Society for Sedimentary Geology, 1957: 124-178. [4] Baker P A, Kastner M. Constraints on the formation of sedimentary dolomite[J]. Science, 1981, 213(4504): 214-216. [5] Land L S. Failure to precipitate dolomite at 25℃ from dilute solution despite 1000-fold oversaturation after 32 years[J]. Aquatic Geochemistry, 1998, 4(3): 361-368. [6] Lippmann F. Crystal chemistry of sedimentary carbonate minerals[M]//Lippmann F. Sedimentary carbonate minerals. Berlin: Springer, 1973: 5-96. [7] Lippmann F. Stable and metastable solubility diagrams for the system CaCO3-MgCO3-H2O at ordinary temperature[J]. Bulletin de Minéralogie, 1982, 105(3): 273-279. [8] Warren J. Dolomite: Occurrence, evolution and economically important associations[J]. Earth-Science Reviews, 2000, 52(1/2/3): 1-81. [9] 李波,颜佳新,刘喜停,等. 白云岩有机成因模式:机制、进展与意义[J]. 古地理学报,2010,12(6):699-710. Li Bo, Yan Jiaxin, Liu Xiting, et al. The organogenic dolomite model: Mechanism, progress and significance[J]. Journal of Palaeogeography, 2010, 12(6): 699-710. [10] Machel H G, Mountjoy E W. Chemistry and environments of dolomitization: A reappraisal[J]. Earth-Science Reviews, 1986, 23(3): 175-222. [11] Kim J, Kimura Y, Puchala B, et al. Dissolution enables dolomite crystal growth near ambient conditions[J]. Science, 2023, 382(6673): 915-920. [12] 甯濛,黄康俊,沈冰. 镁同位素在“白云岩问题”研究中的应用及进展[J]. 岩石学报,2018,34(12):3690-3708. Ning Meng, Huang Kangjun, Shen Bing. Applications and advances of the magnesium isotope on the'dolomite problem'[J]. Acta Petrologica Sinica, 2018, 34(12): 3690-3708. [13] Gaines A M. Protodolomite redefined[J]. Journal of Sedimentary Research, 1977, 47(2): 543-546. [14] Vasconcelos C, McKenzie J A, Bernasconi S, et al. Microbial mediation as a possible mechanism for natural dolomite formation at low temperatures[J]. Nature, 1995, 377(6546): 220-222. [15] Vasconcelos C, McKenzie J A. Microbial mediation of modern dolomite precipitation and diagenesis under anoxic conditions (Lagoa Vermelha, Rio de Janeiro, Brazil)[J]. Journal of Sedimentary Research, 1997, 67(3): 378-390. [16] Sánchez-Román M, McKenzie J A, Wagener A d L R, et al. Presence of sulfate does not inhibit low-temperature dolomite precipitation[J]. Earth and Planetary Science Letters, 2009, 285(1/2): 131-139. [17] Qiu X, Yao Y C, Wang H M, et al. Halophilic archaea mediate the formation of proto-dolomite in solutions with various sulfate concentrations and salinities[J]. Frontiers in Microbiology, 2019, 10: 480. [18] 由雪莲,孙枢,朱井泉,等. 微生物白云岩模式研究进展[J]. 地学前缘,2011,18(4):52-64. You Xuelian, Sun Shu, Zhu Jingquan, et al. Progress in the study of microbial dolomite model[J]. Earth Science Frontiers, 2011, 18(4): 52-64. [19] Petrash D A, Bialik O M, Bontognali T R R, et al. Microbially catalyzed dolomite formation: From near-surface to burial[J]. Earth-Science Reviews, 2017, 171: 558-582. [20] 许杨阳,刘邓,于娜,等. 微生物(有机)白云石成因模式研究进展与思考[J]. 地球科学,2018,43(增刊1):63-70. Xu Yangyang, Liu Deng, Yu Na, et al. Advance and review on microbial/organogenic dolomite model[J]. Earth Science, 2018, 43(Suppl.1): 63-70. [21] Krause S, Liebetrau V, Gorb S, et al. Microbial nucleation of Mg-rich dolomite in exopolymeric substances under anoxic modern seawater salinity: New insight into an old enigma[J]. Geology, 2012, 40(7): 587-590. [22] Kenward P A, Fowle D A, Goldstein R H, et al. Ordered low-temperature dolomite mediated by carboxyl-group density of microbial cell walls[J]. AAPG Bulletin, 2013, 97(11): 2113-2125. [23] Zhang F F, Xu H F, Konishi H, et al. Dissolved sulfide-catalyzed precipitation of disordered dolomite: Implications for the formation mechanism of sedimentary dolomite[J]. Geochimica et Cosmochimica Acta, 2012, 97: 148-165. [24] Zhang F F, Xu H F, Konishi H, et al. Polysaccharide-catalyzed nucleation and growth of disordered dolomite: A potential precursor of sedimentary dolomite[J]. American Mineralogist, 2012, 97(4): 556-567. [25] Liu D, Xu Y Y, Papineau D, et al. Experimental evidence for abiotic formation of low-temperature proto-dolomite facilitated by clay minerals[J]. Geochimica et Cosmochimica Acta, 2019, 247: 83-95. [26] Fang Y H, Xu H F. Dissolved silica-catalyzed disordered dolomite precipitation[J]. American Mineralogist, 2022, 107(3): 443-452. [27] Fang Y H, Zhang F F, Farfan G A, et al. Low-temperature synthesis of disordered dolomite and high-magnesium calcite in ethanol-water solutions: The solvation effect and implications[J]. ACS Omega, 2022, 7(1): 281-292. [28] Meng R R, Han Z Z, Gao X, et al. Dissolved ammonia catalyzes proto-dolomite precipitation at Earth surface temperature[J]. Earth and Planetary Science Letters, 2024, 646: 119012. [29] Han Z Z, Li J, Zhao Y Y, et al. Dissolved Mn2+ promotes microbially-catalyzed protodolomite precipitation in brackish oxidized water[J]. Chemical Geology, 2024, 650: 121986. [30] Chen T, Qiu X, Liu D, et al. Dissolved silicon as a beneficial factor for biomineralization of disordered dolomite by a halophilic cyanobacterium[J]. Chemical Geology, 2024, 670: 122435. [31] Vandeginste V, Snell O, Hall M R, et al. Acceleration of dolomitization by zinc in saline waters[J]. Nature Communications, 2019, 10(1): 1851. [32] Hardie L A. Dolomitization: A critical view of some current views[J]. Journal of Sedimentary Research, 1987, 57(1): 166-183. [33] Kastner M. Sedimentology: Control of dolomite formation[J]. Nature, 1984, 311(5985): 410-411. [34] Liebermann O. Synthesis of dolomite[J]. Nature, 1967, 213(5073): 241-245. [35] Morrow D W, Ricketts B D. Chemical controls on the precipitation of mineral analogues of dolomite: The sulfate enigma[J]. Geology, 1986, 14(5): 408-410. [36] Morrow D W, Ricketts B D. Experimental investigation of sulfate inhibition of dolomite and its mineral analogues[M]//Shukla V, Baker P A. Sedimentology and geochemistry of dolostones. SEPM Society for Sedimentary Geology, 1988: 27-38. [37] Slaughter M, Hill R J. The influence of organic matter in organogenic dolomitization[J]. Journal of Sedimentary Research, 1991, 61(2): 296-303. [38] Compton J S. Degree of supersaturation and precipitation of organogenic dolomite[J]. Geology, 1988, 16(4): 318-321. [39] Brady P V, Krumhansl J L, Papenguth H W. Surface complexation clues to dolomite growth[J]. Geochimica et Cosmochimica Acta, 1996, 60(4): 727-731. [40] Burns S J, Mckenzie J A, Vasconcelos C. Dolomite formation and biogeochemical cycles in the Phanerozoic[J]. Sedimentology, 2000, 47(Suppl.1): 49-61. [41] Holland H D, Zimmermann H. The dolomite problem revisited[J]. International Geology Review, 2000, 42(6): 481-490. [42] Kah L C, Lyons T W, Frank T D. Low marine sulphate and protracted oxygenation of the Proterozoic biosphere[J]. Nature, 2004, 431(7010): 834-838. [43] Li M T, Song H J, Algeo T J, et al. A dolomitization event at the oceanic chemocline during the Permian-Triassic transition[J]. Geology, 2018, 46(12): 1043-1046. [44] Li M T, Wignall P B, Dai X, et al. Phanerozoic variation in dolomite abundance linked to oceanic anoxia[J]. Geology, 2021, 49(6): 698-702. [45] Gregg J M, Kaczmarek S E, Bish D L, et al. A dolomitization event at the oceanic chemocline during the Permian-Triassic transition: Comment[J]. Geology, 2019, 47(7): e467. [46] Rivers J M, Ryan B H, Manche C J, et al. Phanerozoic variation in dolomite abundance linked to oceanic anoxia: Comment[J]. Geology, 2021, 49(9): e534. [47] Frear G L, Johnston J. The solubility of calcium carbonate (calcite) in certain aqueous solutions at 25°[J]. Journal of the American Chemical Society, 1929, 51(7): 2082-2093. [48] Busenberg E, Plummer L N. Kinetic and thermodynamic factors controlling the distribution of SO4 2- and Na+ in calcites and selected aragonites[J]. Geochimica et Cosmochimica Acta, 1985, 49(3): 713-725. [49] Wang X L, Chou I M, Hu W X, et al. In situ observations of liquid–liquid phase separation in aqueous MgSO4 solutions: Geological and geochemical implications[J]. Geochimica et Cosmochimica Acta, 2013, 103: 1-10. [50] Wang X L, Chou I M, Hu W X, et al. Kinetic inhibition of dolomite precipitation: Insights from Raman spectroscopy of Mg2+-SO4 2- ion pairing in MgSO4/MgCl2/NaCl solutions at temperatures of 25 to 200℃[J]. Chemical Geology, 2016, 435: 10-21. [51] Baker P A, Burns S J. Occurrence and formation of dolomite in organic-rich continental margin sediments[J]. AAPG Bulletin, 1985, 69(11): 1917-1930. [52] McCaffrey M A, Lazar B, Holland H D. The evaporation path of seawater and the coprecipitation of Br (super-) and K (super+) with halite[J]. Journal of Sedimentary Research, 1987, 57(5): 928-937. [53] Akilan C, Rohman N, Hefter G, et al. Temperature effects on ion association and hydration in MgSO4 by dielectric spectroscopy[J]. ChemPhysChem, 2006, 7(11): 2319-2330. [54] Liu D, Fan Q G, Papineau D, et al. Precipitation of protodolomite facilitated by sulfate-reducing bacteria: The role of capsule extracellular polymeric substances[J]. Chemical Geology, 2020, 533: 119415. [55] Eigen M, Tamm U K. Schallabsorption in elektrolytlösungen als folge chemischer relaxation II. Meßergebnisse und relaxationsmechanismen für 2—2-wertige elektrolyte[J]. Zeitschrift für Elektrochemie, Berichte der Bunsengesellschaft für physikalische Chemie, 1962, 66(2): 107-121. [56] Rivadeneyra M A, Delgado R, Delgado G, et al. Precipitation of carbonates by Bacillus sp. isolated from saline soils[J]. Geomicrobiology Journal, 1993, 11(3/4): 175-184. [57] Rivadeneyra M A, Delgado G, Soriano M, et al. Precipitation of carbonates by Nesterenkonia halobia in liquid media[J]. Chemosphere, 2000, 41(4): 617-624. [58] Rivadeneyra M A, Párraga J, Delgado R, et al. Biomineralization of carbonates by halobacillus trueperi in solid and liquid media with different salinities[J]. FEMS Microbiology Ecology, 2004, 48(1): 39-46. [59] Jones B F. The hydrology and mineralogy of deep springs lake, Inyo county, California[R]. Reston: USGS, 1965: 56. [60] Jagniecki E A, Berg M D V, Boyd E S, et al. Sulfate-rich spring seeps and seasonal formation of terraced, crystalline mirabilite mounds along the shores of Great Salt Lake, Utah: Hydrologic and chemical expression during declining lake elevation[J]. Chemical Geology, 2023, 636: 121650. [61] Engstrom D R, Nelson S R. Paleosalinity from trace metals in fossil ostracodes compared with observational records at Devils Lake, North Dakota, USA[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1991, 83(4): 295-312. [62] Last W M, Deleqiat J, Greengrass K, et al. Re-examination of the recent history of meromictic Waldsea Lake, Saskatchewan, Canada[J]. Sedimentary Geology, 2002, 148(1/2): 147-160. [63] Lyons W B, Hines M E, Last W M, et al. Sulfate reduction rates in microbial mat sediments of differing chemistries: Implications for organic carbon preservation in saline lakes[M]//Renaut R W, Last W M. Sedimentology and geochemistry of modern and ancient saline lakes models. SEPM Society for Sedimentary Geology, 1994: 13-20. [64] Strang D, Aherne J, Shaw P D. The hydrochemistry of high-elevation lakes in the Georgia Basin, British Columbia[J]. Journal of Limnology, 2010, 69(1s): 56-66. [65] Wright D T, Wacey D. Precipitation of dolomite using sulphate‐reducing bacteria from the Coorong region, South Australia: Significance and implications[J]. Sedimentology, 2005, 52(5): 987-1008. [66] Jacobson G, Jankowski J. Groundwater-discharge processes at a central Australian playa[J]. Journal of Hydrology, 1989, 105(3/4): 275-295. [67] Pirlo M C, Giblin A M. Application of groundwater-mineral equilibrium calculationsto geochemical exploration for sediment-hosted uranium: Observations from the Frome Embayment, South Australia[J]. Geochemistry: Exploration, Environment, Analysis, 2004, 4(2): 113-127. [68] Tweed S, Leblanc M, Cartwright I, et al. Arid zone groundwater recharge and salinisation processes;an example from the Lake Eyre Basin, Australia[J]. Journal of Hydrology, 2011, 408(3/4): 257-275. [69] Papineau M, Haemmerli J. Changes in water quality in the Laflamme Lake watershed area, Canada[J]. Water, Air, and Soil Pollution, 1992, 61(1): 95-105. [70] Simon B, Kucserka T, Anda A. Investigation of Salix alba and Populus tremula leaf litter decomposition in the area of Lake Balaton and Kis-Balaton Wetland[J]. Acta Agraria Debreceniensis, 2018, 74: 159-162. [71] Lopez P L, Mandado J M. Experimental evaporation of superficial brines from continental playa-lake systems located in Central Ebro Basin (northeast Spain)[M]//Schreiber B C, Lugli S, Bąbel M. Evaporites through space and time. London: Geological Society of London, 2007: 143-154. [72] Hidalgo M C, Cruz-Sanjulián J. Groundwater composition, hydrochemical evolution and mass transfer in a regional detrital aquifer (Baza Basin, southern Spain)[J]. Applied Geochemistry, 2001, 16(7/8): 745-758. [73] Camur M Z, Mutlu H. Major-ion geochemistry and mineralogy of the Salt Lake (Tuz Gölü) Basin, Turkey[J]. Chemical Geology, 1996, 127(4): 313-329. [74] Kuşcu M, Şener Ş, Tuncay E B. Recharge sources and hydro geochemical evaluations of Na2SO4 deposits in the Acıgöl Lake (Denizli, Turkey)[J]. Journal of African Earth Sciences, 2017, 134: 265-275. [75] Reimer A, Landmann G, Kempe S. Lake Van, eastern Anatolia, hydrochemistry and history[J]. Aquatic Geochemistry, 2009, 15(1): 195-222. [76] Alipour S. Hydrogeochemistry of seasonal variation of Urmia salt lake, Iran[J]. Saline Systems, 2006, 2(1): 9. [77] Dzhetimov M, Andasbayev E, Esengabylov I, et al. Physical and chemical research of processes of salt formation in the water of Balkhash lake[C]//CBU international conference on integration and innovation in science and education. Prague: CBU, 2013: 400-411. [78] Borzenko S V, Zamana L V, Usmanova L I. Basic formation mechanisms of Lake Doroninskoye soda water, East Siberia, Russia[J]. Acta Geochimica, 2018, 37(4): 546-558. [79] Liu D, Yu N, Papineau D, et al. The catalytic role of planktonic aerobic heterotrophic bacteria in protodolomite formation: Results from Lake Jibuhulangtu Nuur, Inner Mongolia, China[J]. Geochimica et Cosmochimica Acta, 2019, 263: 31-49. [80] Fan Q S, Lowenstein T K, Wei H C, et al. Sr isotope and major ion compositional evidence for formation of Qarhan Salt Lake, western China[J]. Chemical Geology, 2018, 497: 128-145. [81] Cioni R, Fanelli G, Guidi M, et al. Lake Bogoria hot springs (Kenya): Geochemical features and geothermal implications[J]. Journal of Volcanology and Geothermal Research, 1992, 50(3): 231-246. [82] Yan J P, Hinderer M, Einsele G. Geochemical evolution of closed-basin lakes: General model and application to Lakes Qinghai and Turkana[J]. Sedimentary Geology, 2002, 148(1/2): 105-122. [83] Hategekimana F, Ndikuryayo J D, Habimana E, et al. Lake Kivu water chemistry variation with depth over time, northwestern Rwanda[J]. Rwanda Journal of Engineering, Science, Technology and Environment, 2020, 3(1): 1-20. [84] Cheng J R, Meng X Q, Zhang E L, et al. An Early Holocene primary dolomite layer of abiotic origin in Lake Sayram, Central Asia[J]. Geophysical Research Letters, 2021, 48(23): e2021GL096309. [85] Warthmann R, van Lith Y, Vasconcelos C, et al. Bacterially induced dolomite precipitation in anoxic culture experiments[J]. Geology, 2000, 28(12): 1091-1094. [86] van Lith Y, Warthmann R, Vasconcelos C, et al. Sulphate‐reducing bacteria induce low‐temperature Ca‐dolomite and high Mg‐calcite formation[J]. Geobiology, 2003, 1(1): 71-79. [87] Corzo A, Luzon A, Mayayo M J, et al. Carbonate mineralogy along a biogeochemical gradient in recent lacustrine sediments of Gallocanta Lake (Spain)[J]. Geomicrobiology Journal, 2005, 22(6): 283-298. [88] Brennan S T, Lowenstein T K, Cendón D I. The major-ion composition of Cenozoic seawater: The past 36 million years from fluid inclusions in marine halite[J]. American Journal of Science, 2013, 313(8): 713-775. [89] Bathurst R G C. Carbonate sediments and their diagenesis[M]. Amsterdam: Elsevier, 1972: 1-658. [90] Lerman A. Lakes: Chemistry, geology, physics[M]. New York: Springer, 1978: 237-289. [91] Nesbitt H W. The study of some mineral-aqueous solution interactions[D]. Baltimore: Johns PapineauHopkins University, 1974: 173. [92] Siegel F R. Factors influencing the precipitation of dolomitic carbonates[J]. Bulletin (Kansas Geological Survey), 1961(152): 129-158. [93] Deng S C, Dong H L, Lv G, et al. Microbial dolomite precipitation using sulfate reducing and halophilic bacteria: Results from Qinghai Lake, Tibetan Plateau, NW China[J]. Chemical Geology, 2010, 278(34): 151-159. [94] Wells A J. Recent dolomite in the Persian gulf[J]. Nature, 1962, 194(4825): 274-275. [95] Chilingar G V, Bissell H J. Formation of dolomite in sulfate-chloride solutions[J]. Journal of Sedimentary Research, 1963, 33(3): 801-803. [96] Hsü K J, Siegenthaler C. Preliminary experiments on hydrodynamic movement induced by evaporation and their bearing on the dolomite problem[J]. Sedimentology, 1969, 12(1/2): 11-25. [97] Illing L V, Wells A J, Taylor J C M. Penecontemporary dolomite in the Persian gulf[M]//Pray L C, Murray R C. Dolomitization and limestone diagenesis. Tulsa: SEPM Society for Sedimentary Geology, 1965: 89-111. [98] Butler G P. Modern evaporite deposition and geochemistry of coexisting brines, the sabkha, Trucial Coast, Arabian Gulf[J]. Journal of Sedimentary Research, 1969, 39(1): 70-89. [99] Brauchli M, McKenzie J A, Strohmenger C J, et al. The importance of microbial mats for dolomite formation in the Dohat Faishakh sabkha, Qatar[J]. Carbonates and Evaporites, 2016, 31(3): 339-345. [100] DiLoreto Z A, Bontognali T R R, Al Disi Z A, et al. Microbial community composition and dolomite formation in the hypersaline microbial mats of the Khor Al-Adaid sabkhas, Qatar[J]. Extremophiles, 2019, 23(2): 201-218. [101] 袁鑫鹏,刘建波. 回流渗透模式白云岩研究历史与进展[J]. 古地理学报,2012,14(2):219-228. Yuan Xinpeng, Liu Jianbo. Research history and progress on reflux seepage dolostone[J]. Journal of Palaeogeography, 2012, 14(2): 219-228. [102] 朱光有,李茜. 白云岩成因类型与研究方法进展[J]. 石油学报,2023,44(7):1167-1190. Zhu Guangyou, Li Xi. Progress in genetic types and research methods of dolomite[J]. Acta Petrolei Sinica, 2023, 44(7): 1167-1190. [103] 王金艺,金振奎. 微生物白云岩形成机理、识别标志及存在的问题[J]. 沉积学报,2022,40(2):350-359. Wang Jinyi, Jin Zhenkui. Formation mechanism, identification markers, and questions regarding microbial dolomite[J]. Acta Sedimentologica Sinica, 2022, 40(2): 350-359. [104] Perri E, Tucker M. Bacterial fossils and microbial dolomite in Triassic stromatolites[J]. Geology, 2007, 35(3): 207-210. [105] 王小林,胡文瑄,陈琪,等. 塔里木盆地柯坪地区上震旦统藻白云岩特征及其成因机理[J]. 地质学报,2010,84(10):1479-1494. Wang Xiaolin, Hu Wenxuan, Chen Qi, et al. Characteristics and formation mechanism of Upper Sinian algal dolomite at the Kalpin area, Tarim Basin, NW China[J]. Acta Geologica Sinica, 2010, 84(10): 1479-1494. [106] 由雪莲,孙枢,朱井泉. 塔里木盆地中上寒武统叠层石白云岩中微生物矿化组构特征及其成因意义[J]. 中国科学:地球科学,2014,44(8):1777-1790. You Xuelian, Sun Shu, Zhu Jingquan. Significance of fossilized microbes from the Cambrian stromatolites in the Tarim Basin, Northwest China[J]. Science China Earth Sciences, 2014, 44(8): 1777-1790. [107] 胡文瑄,朱井泉,王小林,等. 塔里木盆地柯坪地区寒武系微生物白云岩特征、成因及意义[J]. 石油与天然气地质,2014,35(6):860-869. Hu Wenxuan, Zhu Jingquan, Wang Xiaolin. Characteristics, origin and geological implications of the Cambrian microbial dolomite in Keping area, Tarim Basin[J]. Oil & Gas Geology, 2014, 35(6): 860-869. [108] Liu S A, Liu P P, Lv Y W, et al. Cu and Zn isotope fractionation during oceanic alteration: Implications for oceanic Cu and Zn cycles[J]. Geochimica et Cosmochimica Acta, 2019, 257: 191-205. [109] Campbell K A. Hydrocarbon seep and hydrothermal vent paleoenvironments and paleontology: Past developments and future research directions[J]. Palaeogeography, Palaeo-climatology, Palaeoecology, 2006, 232(2/3/4): 362-407. [110] Orphan V J, House C H, Hinrichs K U, et al. Methane-consuming archaea revealed by directly coupled isotopic and phylogenetic analysis[J]. Science, 2001, 293(5529): 484-487. [111] Peckmann J, Thiel V. Carbon cycling at ancient methane–seeps[J]. Chemical Geology, 2004, 205(3/4): 443-467. [112] Ritger S, Carson B, Suess E. Methane-derived authigenic carbonates formed by subduction-induced pore-water expulsion along the Oregon/Washington margin[J]. Geological Society of America Bulletin, 1987, 98(2): 147-156. [113] Aloisi G, Gloter A, Krüger M, et al. Nucleation of calcium carbonate on bacterial nanoglobules[J]. Geology, 2006, 34(12): 1017-1020. [114] Rivadeneyra M A, Delgado R, Párraga J, et al. Precipitation of minerals by 22 species of moderately halophilic bacteria in artificial marine salts media: Influence of salt concentration[J]. Folia Microbiologica, 2006, 51(5): 445-453. [115] Tong H P, Feng D, Peckmann J, et al. Environments favoring dolomite formation at cold seeps: A case study from the gulf of Mexico[J]. Chemical Geology, 2019, 518: 9-18. [116] 张亦凡. 类白云石结构碳酸盐的低温合成及微结构研究[D]. 合肥:中国科学技术大学,2020:1-151. Zhang Yifan. Study on the low-temperature syntheses and microstructure of dolomite-analogous carbonates[D]. Hefei: University of Science and Technology of China, 2020: 1-151. [117] Farkas L, Bolzenius B H, Will G. Powder diffraction data and unit cell of kutnohorite[J]. Powder Diffraction, 1988, 3(3): 172-174. [118] Beran A, Zemann J. Refinement and comparison of the crystal structures of a dolomite and of an Fe-rich ankerite[J]. Tschermaks Mineralogische und Petrographische Mitteilungen, 1977, 24(4): 279-286. [119] Rosenberg P E, Champness P E. Zincian dolomites and associated carbonates from the Waryński Mine, Poland: An AEM investigation[J]. American Mineralogist, 1989, 74(3/4): 461-465. [120] Lippmann F. Syntheses of BaMg (CO3)2 (norsethite) at 20 ℃ and the formation of dolomite in sediments[M]//Müller G, Friedman G M. Recent developments in carbonate sedimentology in Central Europe. Berlin: Springer, 1968: 33-37. [121] Effenberger H, Zemann J. Single crystal X-ray investigation of norsethite, BaMg(CO3)2: One more mineral with an aplanar carbonate group[J]. Zeitschrift für Kristallographie-Crystalline Materials, 1985, 171(3/4): 275-280. [122] Secco L, Lavina B. Crystal chemistry of two natural magmatic norsethites, BaMg(CO3)2, from anmg-carbonatite of the alkaline carbonatitic complex of Tapira (Se Brazil)[J]. Neues Jahrbuch für Mineralogie Monatshefte, 1999(2): 87-96. [123] Effenberger H, Pippinger T, Libowitzky E, et al. Synthetic norsethite, BaMg(CO3)2: Revised crystal structure, thermal behaviour and displacive phase transition[J]. Mineralogical Magazine, 2014, 78(7): 1589-1611. [124] Lippmann F. PbMg(CO3)2, ein neues rhomboedrisches doppelcarbonat[J]. Naturwissenschaften, 1966, 53(24): 701. [125] Goldsmith J R. Cadmium dolomite and the system CdCO3-MgCO3 [J]. The Journal of Geology, 1972, 80(5): 617-626. [126] Froese E. A note on strontium magnesium carbonate[J]. The Canadian Mineralogist, 1967, 9(1): 65-70. [127] Bragg W L. The analysis of crystals by the X-ray spectrometer[J]. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences, 1914, 89(613): 468-489. [128] Kharaka Y K, Barnes I. SOLMNEQ. Solution-mineral equilibrium computations. [Equilibrium distribution of aqueous species in natural waters at 0 to 350/sup 0/][D]. Berkeley: University of California, 1973. [129] Dean J A. Lange's handbook of chemistry[M]. New York: McGraw-Hill, 1999. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1162
- HTML全文浏览量: 149
- PDF下载量: 146
- 被引次数: 0

