

 下载:
下载:



-
二叠纪是古生代最后一个纪,在华南发育有比较连续的地层记录。该时期地壳运动趋于活跃,全球范围内一系列板块的碰撞导致地史中著名的联合古陆在二叠纪末期基本形成。这种全球古构造、古地理的巨变,造成了气候带的明显分异和生物界的重要变革[1-2]。在二叠纪末期发生的生物灭绝被众多学者研究,但此次灭绝事件的驱动因素仍在争论中[3]。作为史上五次大灭绝最为严重的一次,陆地系统[4]和海洋系统[5]均遭到严重地破坏,其中底栖生物群遭到重创,几近灭绝。正因如此,中国华南的乐平统和二叠系—三叠系界线附近是研究程度最高的地层,其中的地质年代学、生物地层、化学地层综合框架等均是全球对比的标准[6-10]。
在二叠纪早期,冰期的影响占据着影响生物种群演化的主要位置,多变的气候是限制生物发展的主要因素[11]。到二叠纪中期,由季风所产生的气候改变及水体变化[12-13]从另一方面影响了生物群落的结构。对于二叠纪末期生物绝灭的过程已也有诸多认识[4,14-16],其中起到生物绝灭的推动因素包含海洋缺氧[17-19],风化作用增强[20-21] 和高温[22] 等因素。
腕足类是二叠纪广泛存在的底栖滤食性海洋生物群,可以在不同的环境下做出生态响应,能很好的反映出当时的古环境变化,因此有助于古环境重建。一些学者已经对全球二叠纪腕足类宏演化过程进行了恢复并进行探讨,如Shen et al.[23]对整个华南石炭纪晚期至三叠纪早期的腕足类数据进行了多样性时间变化的研究。但是有关腕足类的多样性的控制因素,环境变化及响应机制仍缺少研究。本文选择中扬子区新滩剖面腕足类为研究对象,通过创建亚丁斯克阶至长兴阶的连续生物多样性曲线,根据海平面变化及C、O同位素[23]变化分析控制因素,得到了高分辨率研究的结果并与华南数据进行比对分析,旨在恢复以腕足类视角所体现到的古环境的变化。
-
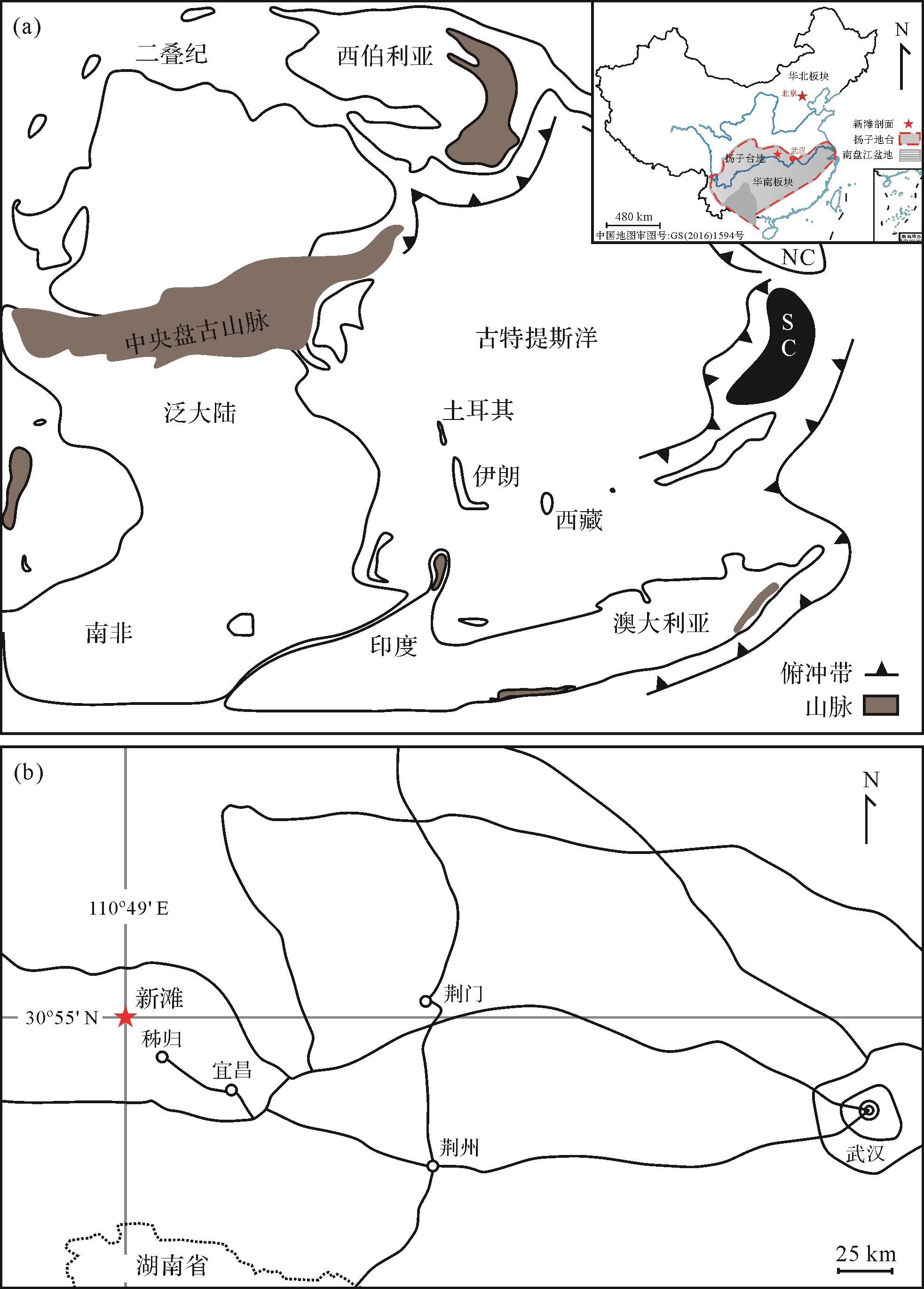
华南中—上扬子板块是研究二叠纪地层的经典区域之一,受控于泛大陆的聚合和裂解活动[24],期间受到3个阶段的大规模火山活动影响:1)早二叠世塔里木大火成岩省[25];2)中二叠晚期的峨眉山大火成岩省[26-27];3)P-T(Permian-Triassic)之交广泛分布于华南地区的多层火山灰为特征的酸性火山作用[28-29]及历史时期最大规模的西伯利亚大火成岩省[30-31]。生物演化经历了石炭纪晚期—乌拉尔世早期的整体稳定繁荣阶段、瓜德鲁普世的波动阶段以及P-T之交的显生宙最大规模生物灭绝阶段。气候经历了冰期转温室转低温转炎热的环境转换[32]。华南板块内沉积相分异较大[33],二叠纪期间主要包括浅水台地,斜坡陆棚,深水盆地(图1)。

新滩剖面位于湖北省宜昌市秭归县三峡大坝上游约20 km处。区域地层从下至上为石炭统黄龙组顶部,下二叠统马鞍山段(梁山组)及栖霞组,中二叠统茅口组,上二叠统吴家坪组与长兴组,三叠统大冶组底部。栖霞组与马鞍山段总厚为212.4 m,其底部以约2.8 m马鞍山段(梁山组)黑色泥岩为显著特征与黄龙组灰岩为平行不整合,下部多发育生屑灰岩(䗴类,有孔虫类,珊瑚类,腕足类,介形类)且富含藻屑,中部发育带状及团块状燧石,上部发育黑色瘤状灰岩。茅口组总厚为178.25 m,其底部发育灰岩夹薄层白云岩,中下部发育燧石结核及生物碎屑(䗴类,有孔虫类,腕足类),中部多见藻类发育,上部以生屑灰岩发育为主,其间夹薄层白云岩且含少量燧石结核,最顶部为1 m茅口组王坡页岩段。吴家坪组总厚46.31 m,整体以生屑灰岩(䗴类,有孔虫类,珊瑚类,腕足类)发育为主,中部含团块及串珠状燧石。长兴组总厚为37.7 m,整体发育以生屑灰岩(䗴类,有孔虫类,珊瑚类,腕足类)为特征,顶部发育藻类,含少量燧石结核。大冶群组最底部有一层薄层泥灰岩,主体为黑色泥灰岩沉积,富含黄铁矿。
-
根据前人的研究,选取出露完整的新滩剖面开展腕足类研究。剖面的地层、岩性及化石信息主要来自冯少南等[35]编著的专著《长江三峡地区生物地层学(3)晚古生代分册》的专著。化石数据采用来自上述专著及Rong et al.[36]编著的专著Phanerozoic Brachiopod Genera of China,在进行腕足种类的校对之后,得到腕足类的地层分布。由于华南上扬子区内䗴类较为发育,且牙形石发育不完全,遂将剖面资料根据䗴带及中国二叠纪最新框架[37]进行时间划分。
按照种级和属级计算简单多样性N tot:
(1) 按照种级和属级估算平均多样性N emsd [38]:
(2) 绘制腕足类多样性曲线及趋势投影,并与华南腕足多样性[23]进行对比分析。其中,FL表示该化石首现层位位于时间段内,同时其末现层位也位于该时间段内,称之为单延限分子;bL代表首现层位位于时间段底界之下,末现层位位于时间段内的化石类型,Ft代表首现层位位于时间段内,末现层位位于时间段顶界之上的化石类型,这两种类型统称为半贯穿分子;bt代表化石首现和末现层位均位于时间段界线之外的化石类型,称之为贯穿分子[38]。
-
峡区二叠纪的腕足类相当丰富,从剖面产出共计48属92种(图2),根据其组合特征可自下而上分为五个组合。在以下各节中,以自下而上的地层顺序描述腕足组合,其中具代表性的腕足种类均参考自Phanerozoic Brachiopod Genera of China专著[36]第二卷及尚庆华等[39]的研究结果。

图 2 中扬子新滩地区二叠纪腕足类种级延限
Figure 2. The species⁃level range of Permian brachiopods in Xintan area, Middle Yangtze
-
栖霞组底部的马鞍山段(梁山组)对应亚丁克斯阶,其出产Orthotichia chekiangensis-Ogbinia hexaspinosa组合,Ogbinia hexaspinosa为鄂西栖霞组中下部典型代表之一。Tyloplecta richthofeni、O. chekiangensis等广布于华南同期地层。同时Orthotichia也是西特提斯区腕足类的优势群。
-
栖霞组中上部对应空谷阶及罗德阶底部,其出产Cryptospirifer semiplicatus-Tyloplecta nankingensis组合,常见分子有Cryptospirifer semiplicatus、C. omeishanensis,C. minor、Tyloplecta nankingensis,Monticulifera sinensis、Chilianshania chilianshanensis及Urushtenoidea chaoi等。Cryptospirifer这种巨大的冷水隐石燕贝类的大量富集为本组合首要特点,并常见于亚美尼亚,伊朗,土耳其的上乌拉尔统至下瓜德鲁普统。
-
茅口组对应罗德阶、沃德阶及卡匹墩阶,主要出产Neoplicatifera huangi-Monticulifera hunanensis组合,该组合主要代表了茅口组灰岩段腕足类动物群,其晚期主要出产Neoplicatifera huangi,是华南早二叠世晚期特征化石之一,属于冈瓦纳冷水动物群。值得一提的是,该组合出产的Orthotichia暖水型动物群在新几内亚二叠纪腕足类动物群[40]中同样被发现,说明该区具有冈瓦纳型冷水动物群和特提斯型暖水动物群过渡性质。Cryptospirifer类和Vedeproductus等仅在华南至土耳其一带出现,构成了独特的腕足动物区。总体生物群面貌以长身贝类和戟贝类为主体。
-
吴家坪组对应吴家坪阶,其出产Tschernyschewia sinensis-Lopingia ruber组合,主产于吴家坪组下部。组合分子除了不少为中二叠世延续分子以外,也有许多新分子出现。其中的Perigeyerella,Waagenites,Enteletes等都是作为华南生物地层对比的重要分子[37]。此外,Squamularia在本区十分丰富,种的分异较为明显。
-
长兴组对应长兴阶,出产Squamularia grandis-Richthofenia sinensis组合。在晚二叠世长兴期,长兴期腕足类动物已经衰退,正形贝类、长身贝类等各类群的属种数量减少;石燕贝类以光面石燕为主。在长兴末期的腕足类主要发生身体结构改变,如隔板增强的Richthofenia,Leptodus等,个体大型化发展的Squamularia grandis,Tyloplecta yangtzeensis等,其中首要分子Sguamularia grandis常见于长兴期。
-
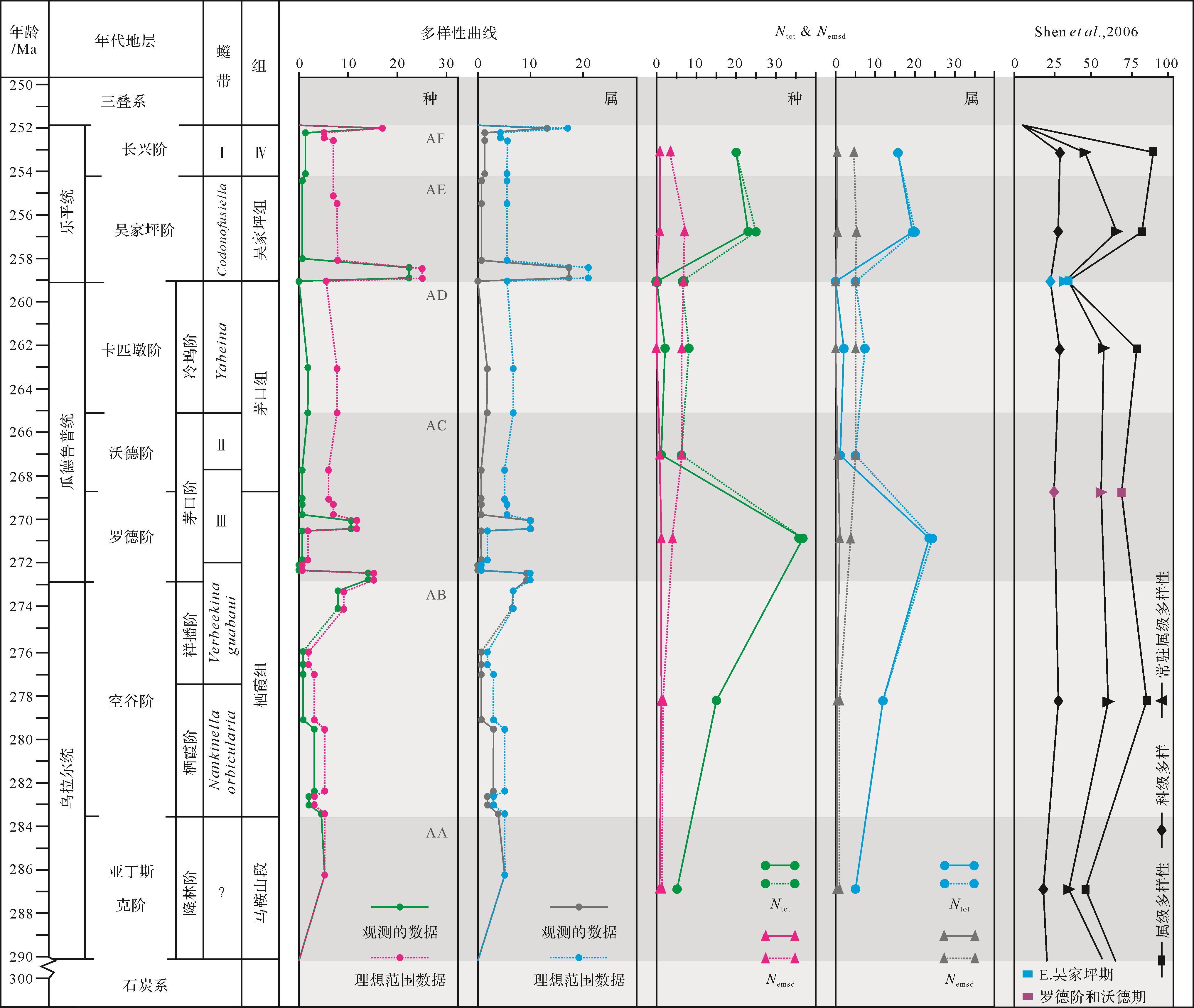
根据观察到的物种多样性分析了新滩区早、中、晚二叠世腕足类的种群变化(图2),及理想范围类种群的变化(图3)。在图3中,多样性曲线可供识别多样性峰值为4个,通过计算N tot、N emsd后,共识别出2个峰值,为了便于与其他数据及前人研究[23]进行比较,按照阶划分为6个区域,其中罗德阶和沃德阶为了与华南研究对应,暂合并为AC一个区域讨论。

图 3 中扬子区腕足类多样性曲线与华南腕足数据对比(I:Palaeofusulina sinensis带;II:Neoschwagerina haydeni带;III:Chusenella conicocylindrica带;IV:长兴组)
Figure 3. Comparison of brachiopod diversity curves in middle Yangtze region and brachiopod data in South China (Fusulines: I. Palaeofusulina sinensis. II. Neoschwagerina haydeni. III. Chusenella conicocylindrica. IV. Changhsingian Formation)
腕足类多样性曲线在经历了AA段的平稳发展后,于AB段早期和中期发生了小幅度的波动,这部分波动使得在N tot结果中,腕足类多样性发展有了上升趋势,这部分上升趋势与华南数据的上升趋势一致。而AB段末中的N tot上升趋势与华南数据呈现相反趋势,腕足类多样性曲线得到了增长,发生了一定规模的辐射,不过此次多样性曲线增长过后却迅速在AC段下降至冰点,并在之后又进行一次小规模增长。这两个峰值之间的变化主要表现在以Cryptospirifer为首的一部分冷水种群消失,以Orthotichia为首的暖水种群出现,以暖水分子与冷水分子共同出现在这一阶段为特征。由于这些分子的高分异度发展,使得N tot明显在AC段出现多样性峰值,但华南属级多样性在AC段呈现出先下降再上升的趋势,与研究结果不同,但华南常驻属级多样性发展与N emsd数据实际表现相对一致。随后多样性曲线在大幅下降之后以略微上升再下降的小幅度波动姿态发展到AD段末期,与华南数据的显示结果相近。之后多样性曲线与N tot曲线于AD&AE交界处下降至冰点,华南数据中也在E.吴家坪期均有了下降趋势,其中属级多样性下降幅度巨大。不过这一阶段从理想范围数据与N emsd的曲线上看,种群的数量变化是较为缓和的,下降幅度仅在二分之一左右。在AE段的结果显示,种群在冰点过后立即出现多样性峰值,其中的腕足类除以Enteletes等为首的新生分子出现以外,其余分子大多是来自上2个层位的延续分子。同时AE段N tot与N emsd曲线中的上升趋势与华南数据所显示的趋势都同为上升趋势。多样性曲线在经历此次峰值之后又回归至AD段的水准。在最后的AF段,位于末期的数据显示,种群的数量又达到了一个新的峰值,之后在极短的时间内发生陡崖式跌落,使得该区域的腕足多样性跌至零点。其中的华南数据中显示,属级多样性为上升趋势再下降,常驻属级多样性则一直为下降趋势。
其中,在N emsd和华南数据中的常驻属级多样性曲线及科级多样性曲线中共同表明,三者曲线并没有剧烈波动,而是一直以相对平稳的方式贯穿整个二叠纪。
-
在中扬子新滩剖面中保存了完好的腕足记录。到目前为止,华南牙形石、䗴类、有孔虫、腕足、珊瑚等生物格架已被记录为中国二叠纪生物地层框架[37]。华南上扬子环境背景包括建立高分辨率海平面曲线[41-42]、碳同位素趋势、牙形石氧同位素趋势[43]及锶同位素比值分析[44]。针对特提斯大区,同样有来自阿曼、西西里和伊朗的基于腕足壳体的碳同位素及氧同位素(图4)[45]。从碳同位素的正偏与负偏可以看出当前区域的生产力变化与营养水平[46-47],氧同位素的正偏意味着温度的升高,冰期的结束等[48],δ 87Sr/δ 86Sr比值的增长可以看出风化作用是否增强及气候演变[44]等。通过这些数据可以得到同一个区域的生物多样性在不同环境下做出了不同的改变,或者假设在全球变暖的情况下,局部区域的生物是否能对全球变暖做出响应变化。N tot曲线代表短延限分子,以化石产出记录为对象、以时间段为单位的编目式物种多样性统计分析,较好体现出生物群辐射及灭绝的强度,方便对比每个时期之间的种群变化趋势,可与华南数据的多样性指数作对比;N emsd曲线代表长延限分子,以常驻腕足类分子为对象,与华南常驻多样性意义基本一致,展示了生物群的整体趋势,理想范围数据为实际观测数据的补充,纠正了因实际化石产出带来的偏差。在环境对比方面,选择了基于观测数据和理想范围类数据的多样性曲线为主进行分析(图4)。在AA段至AB段,Nt ot曲线与华南数据相对一致,整体呈上升趋势,说明该阶段的多样性呈现增多趋势,而多样性曲线并无上升趋势,处于波动阶段,N emsd趋势较为平稳,表明该阶段的上升趋势并非是腕足类发生了辐射引起的,而是整个华南腕足类种群的演变造成了这种结果。在AB段至AC段,N tot出现了峰值,与华南数据呈现相反趋势,通过多样性曲线可以看出,峰值的出现是由于强烈波动与上升趋势共同引起的结果,这说明该时期出现了区域上的独特变化,在区域上产生了辐射,与华南大区产生了分异。在AC至AD段,多样性曲线与N tot曲线在AC段中期出现回落但在AD早期呈现略微上涨趋势,表明研究区与华南大区进入趋同阶段;通过N emsd曲线可以看出,AC至AD的整体阶段是较为平稳的,与华南数据中的常驻多样性较为接近,展示了生物群的整体稳定性。在AD段末,所有曲线都呈现略微的下降趋势,但与同时期的华南数据中属级多样性的大幅度跌落相比,是较为稳定的,其中华南数据中常驻属级多样性发生了小幅跌落,说明整个大区出现了腕足类危机,但研究区未受到过多影响;或者是整个华南数据的置信区间被过分估算,高估了危机程度。最后在AE段至AF段,腕足类在早期出现了大范围辐射之后,进入稳定发展状态,所有的曲线趋势基本一致。其中,AF段中华南属级曲线相对其他曲线偏高是华南乐平动物群的最深入研究[23]导致,因此常驻属级多样性更为可信。
-
在乌拉尔世,腕足类的多样性曲线的发展趋势较为平缓,研究区中主要为Tyloplecta等为首的冷水分子作为观察对象,通过多样性曲线可以看出,腕足类在经历了AA段的平稳发展后,于AB段中部数量下降,最后在AB段末有了明显的增长。但根据N tot结果及华南数据显示(图3),二者曲线在AB段的整体趋势较为平稳,并且渐有上升趋势,对比分析得出研究区冷水分子在AB阶段的下降更可能是由于地层缺失导致。由于石炭纪冰期消融的主幕发生在此阶段,并一直持续到瓜德普鲁统末期,且通过AA及AB阶段的δ 18O数据[45](图4)得出,负偏信号指示了腕足类从乌拉尔世早中期同样处在冷水环境中。而研究剖面中亚丁斯克阶的黑色钙质泥岩,空谷阶的黑色团块燧石及黑色碳质泥岩反应了缺氧的环境。因此较低的温度加上贫瘠的环境有可能是造成多样性曲线平缓的主要原因。
在剖面的栖霞组岩性中,生屑灰岩的生屑种类为䗴类,有孔虫类,珊瑚类,介形类,腕足类。其中䗴类、有孔虫及藻类的大量发育表明浮游生物所处环境较好,说明表层水体开始出现变暖迹象,这与水体分层观点是一致的[49]。在针对牙形石氧同位素的研究中表明[43] ,乌拉尔世中后期δ 18O同样开始出现正偏趋势。所以水体温度的逐渐上升为腕足类的多样性发展提供有利条件,从而在多样性曲线中呈现上升趋势。在新滩剖面中,以Hayasakaia为顶峰代表所出现的皱纹珊瑚中表明,珊瑚均为小型—中小型个体,反映了冷水洋流的存在[50] 。不过需要注意的是,在研究剖面中所体现出的珊瑚物种的分异度往往和腕足类繁盛呈负相关关系,当珊瑚分异度高时,腕足类就相对较低,这种对立关系在当今的红海仍然存在[51] 。关于这一点,除了盐分因素影响以外,珊瑚与腕足的竞争关系主要在于其都是滤食性的,同时腕足幼体对固着底质有一定要求(比如想固着在珊瑚表面就较为困难)。所以,作为底栖生物的腕足类,生存条件受水体环境及生物竞争的双重影响。
-
在乌拉尔世末期至瓜德鲁普世早期阶段,联合古大陆已基本进入联合完成阶段,赤道附近的中央盘古山脉隆起,使位于南北半球的高纬度冰川和赤道之间形成温度梯度[52],同时特提斯洋作为赤道附近的大型暖池,控制着季风的大小及位置[52],使特提斯大区进入温室气候并发育出特殊的季风系统。这种“超级季风”气候使陆地的生物化学风化作用增强[53-54],加速了养分向海洋的补给过程。空谷阶上部的瘤状灰岩表明,洋流出现了增强[55],养分通过洋流输送进海洋可能是该时期腕足辐射的主要原因。研究表明,导致这次辐射的因素与奥陶纪生物大辐射极为相近的[56]。通过分析腕足类多样性曲线发现,曲线进行了两个阶段式的演变。第一段演变于AB末期,结束于AC早期,种的分异度明显大于属的分异度,这样的趋势证明了水体营养结构较好且环境稳定。第二段演变始于AC早中期,结束于AC中期,属级之间出现了变化,以Orthotichia等为首的暖水分子占据了主要演化位置,而Neoplicatifera等冷水分子与Cryptospirifer分别出现了凋零与消失现象。这些分子之间的变化揭示了水体由冷转暖的过程,且该时期δ 18O的升高同样证实了这一过程[43]。通过上述结果及前人的研究[12-13]可以看出,巨型季风不仅改善了水体营养结构,加速了水体循环,甚至对冰川消融也起到了一定作用,导致水体从冷水快速转变为暖水,最终导致腕足类群在相对短时间发生改变。另一方面,该时期发育的白云岩也说明在乌拉尔世末期至瓜德鲁普世早期阶段之间有强烈的蒸发作用存在。蒸发作用会使水体产生强烈的上升流,使洋盆深处的缺氧水被带到上层位置,加上该时期的华夏古陆隆起使得浅海和小洋盆之间的水体交换加快,最终导致腕足动物群在研究区与华南大区同样环境下的不同种群分异。在图3的沉积相中可以看出,整个栖霞组经历了从深水陆棚相到浅水台地相的转变,同时在转变过程中,研究区由于受到季风的影响使得环境更加复杂多变。在两段辐射的中间只观察到湖北特有种Monticulifera及Songzichonetes sanxiaensis,两者身体皆小型化且两壳扁平,不仅更益于适应上述复杂多变的环境,同时也是对上述环境的一种印证。不过二段式发展并没有在华南动物群中体现,这种不同的结果说明了研究区受到了空间限制作用影响,但华南种群体现的衰败也同样体现在冷水种的减少,且内蒙古地区同样有混生现象[57],说明位于中二叠世不稳定系统下环境的一致性。
在同为中扬子板块的四川渡口剖面茅口组的研究中,在冰川融化过程呈现了从间冰期到大冰期的转换[58] 。通过对氧同位素趋势(图4)可以看出,在瓜德普鲁世晚期,δ 18O负偏移的趋势预示了气候的变化[48],这意味着该区域腕足类所在的环境正在变冷,在时间线上与在卡匹墩期末期的Kamura寒冷事件[59]呈现一致性。从剖面的观测来看,茅口组中期AC和AD段多发育含藻类、䗴类及有孔虫等大量生屑灰岩与白云岩,整体沉积环境以浅海台地为主。腕足的多样性在观测数据中接近零点,理想范围数据显示多样性下降一半左右,并一直稳定。通过碳同位素的波动信号表明生产力出现了高频变化,印证了浮游生物大量繁盛,争夺了底栖生物的氧气与养分资源[60],使腕足在实际观察数据中位于低值。但从多样性的理想范围数据的表现来看,腕足类动物群只是不能适应研究区的环境,转移到研究区之外,如当研究区在深水环境转浅水环境再转深水环境过程中,深水生物会发生转移现象。在理想范围数据中,腕足以Squamularia,Araxathyris,Cathaysia为主要研究对象。其中Squamularia个体发育较大,多见于碳酸盐台地当中[61],Cathaysia个体较小,壳体较薄,在华南多发育于硅质岩中见于半深水—深水环境中[62] ,两者说明研究区附近同时存在上述两种环境。在卡匹墩期的Kamura事件中,腕足类群并未受到影响,但珊瑚遭受了强烈影响[63]。从整体上看,研究区的茅口组经历了早期的变暖至末期变冷的一个过程,并在末期出现了一次大的海退事件[58] 。综合上述因素的影响,腕足类多样性在剖面中显现度较低,但延续性并未中断,发展趋势依旧良好。
-
前乐平统危机在剖面中的体现为1 m厚的黏土岩段,AD段末尾中未观测到腕足存在的迹象(图3)。在经历了灭绝第一幕的前乐平统事件及峨眉山火成岩省事件之后,AE段初的腕足多样性在剖面达到二叠纪的峰值,有延续而来的Spinomarginifera,Squamularia,Cathaysia等延续分子,也有Perigeyerella,Waagenites,Enteletes等新生分子出现。腕足种群出现了多元化发展,其中代表性的Tyloplecta为假疹发育的老分子,Squamularia为体型较大的分子,Edriosteges,Spinomaraginifera为壳刺发育的新分子。此次的发展和海平面的快速升高与营养的迅速补给(事件造成了强烈的陆地风化作用,使陆源物质大量进入海洋)有密切的关联[64]。沉积表现也多为硅质岩及藻泥晶灰岩,䗴类及有孔虫等未在剖面位置显现。在峨眉山事件中心,华南腕足类显示种级的灭绝率为87%,但属级灭绝只有30%[23] ,而从剖面上的响应结果来看,大部分辐射中的分子都来源于延续分子的异化,N emsd数据与华南腕足类科级数据中显示,两者的曲线发展方式都是较为平稳的。这说明以往的G-L事件的确是引起生物种群变化的因素之一[65-66],但被过分高估了其破坏能力,至少在腕足中是这样的。
在吴家坪早期到长兴末期,腕足以较稳定的方式发展。其中壳刺类及大型腕足类的发展可能与大量的发育的碳酸盐台地有关[67],而小型化,壳体较薄且壳饰弱化的种存在于深水缺氧和营养贫瘠环境中。这一点与二叠纪初期是一致的,具代表性的Tyloplecta贯穿整个二叠纪。在P-T事件中,研究区内腕足呈现了灭绝的结果,与华南动物群中的的长身贝目、扭月贝目、石燕目和直形贝目等均灭绝[23]呈现出一致性。关于灭绝前AF段的峰值,作者认为是环境出现差异性导致的。在浅海台地环境中,δ 13C出现负漂,通过生产力的下降可以看出区域内的浅水生物群落的消失给了腕足爆发性增长的时机,需要注意的是长兴晚期的变冷事件限制了珊瑚的发展[68],减少了底栖生物的竞争。而深水环境的腕足此时同样存在,这样导致同一区域内存在的多种环境使腕足的种群分异度明显增加。随后的灭绝事件,作者更支持西伯利亚火山的喷发可能是造成环境剧变的诱因这一观点[3] ,同时也是气候变暖,海洋环境恶化的催化因素。大气环境的剧变与大规模海退事件使浅海发生了强烈的破坏作用,海水的酸化与快速升温的观点[69]被广泛接受。在深水中,体系的崩坏甚至早于浅水,这一点从高纬度的硅质海绵崩坏可以体现[70]。洋流是一直存在的,因火山喷发造成缺氧且缺营养的水会极大的限制深水腕足的生态位,通过扩散作用进一步影响到浅海[70]。在浅海中,表层水受到酸化高温[71]等影响,大规模海退事件[72]使生存空间减少。在这种情况下,腕足的生存空间接近阈值,最终导致该区域内腕足的灭绝。
-
在研究区中,腕足类受到区域相变等控制,出现了区域上的特有结果。在研究观察到的腕足分异数据中,AB-AC与AD-AF在环境反映中出现了较高的相似性,二者都产生了腕足类种群变化,每一段都由多种环境(温度变化,海平面变化等)共同驱动,导致一个时期的较高分异度,在N tot及N emsd曲线上呈现出两个峰值的结果。二者都在δ 13C表现中发生了高频的波动,而且都经历了δ 18O的负偏移,同时具有转温室的迹象及洋流影响显著增强[73]。在洋流影响方面,前者洋流的增强表现为使Cryptospirifer等分子在华南至土耳其一带形成独特的腕足区,这是由于底栖固着类生物想要扩散到其它较远区域只能依赖水体的流动[74-75],后者的表现为使Edriosteges等分子为了更好的固着于海底而高度发育壳刺[64]。但这一现象并没有很好的在华南数据中体现出来,仅有一小部分能从Nemsd曲线中看出趋势,如AB段及AD段。而且通过耦合地壳活动发现,这两次事件变化之前都有火山活动[76],与火山活动时间的间隔呈现缩短趋势并且出现了强烈的环境变化,最后在PT事件中出现了高度融合,这部分原因还有待研究。
-
在上述对早—中二叠世的研究中表明,冰期所产生的气候变化影响着研究区腕足的多样性变化。水体的分层及生物间的竞争关系也起到了一定的限制作用。在冰期消融期间,研究区域出现了冷水分子与暖水分子的混合过渡带。温室效应及“巨型季风”使陆源营养物质注入海洋并对腕足类的辐射起到了间接催化作用。在华南构造运动下,研究区的茅口组早期经历了深水陆棚相至浅海台地相的转变,环境产生了不同的变化,使腕足类动物群在针对同一区域的不同环境得到了多元化发展。在晚二叠世,Kamura事件,峨眉山和西伯利亚火山事件相继发生,但通过腕足表现来看,前两者的影响并没有造成灾变级影响,反而西伯利亚火山事件带来了无论是整个生态系统还是局限环境的灾难性破坏,从上至下贯穿整个生态域。腕足类在二叠纪中期与晚期都发生了种群及形态的改变,在区域上呈现了特有的结果,且两个阶段的演变方式和原因都与气候变暖有关,每个阶段峰值之间接近6~8个百万年,每一个峰谷之间的变化都在2个百万年内,但由于展示的仅是该区域腕足类所体现的变化,因此具体的原因还需要通过后续开展对不同生物之间的多样性来研究补充。
Permian Brachiopods in Xintan, Western Hubei:Community evolution model and response to environmental changes
-
摘要:
系统研究了湖北省西部新滩剖面亚丁斯克阶至长兴阶的腕足动物群落演替过程。通过研究区与华南地区及特提斯区域进行比对分析,结合前人对碳、氧同位素的研究结果,分析了二叠纪腕足类群落演替的规律和区域差异。结果表明:除二叠纪末的灭绝事件,在乌拉尔统与瓜德鲁普统界限附近及乐平统中共识别出两次腕足类种群的显著分异度变化,且其演替时间都接近6~8个百万年。前一次体现在冷水种与暖水种的混合过渡现象,后一次体现在个体增大、壳饰发育与个体变小、壳体变薄的不同分化现象。在气候方面,冰期与季风的强弱作为主要因素从温度及养分来源共同影响腕足类的多样性变化。火山喷发事件及板块运动改变腕足类生态环境并造成区域的分化。海平面变化既受气候影响也受地壳运动影响,高频的海平面变化不断改变着腕足类在研究区的生存空间。腕足类在适应不同环境中所发生的壳体形态改变、群落结构演替很好的展示了生物环境对古环境的响应。 Abstract:A systematic study of the succession process of the brachiopod community from Artinskian to Changhsingian at Xintan, western Hubei Province is described. The causes and regional differences within the Permian brachiopod community succession were analyzed by comparative analysis of the study area, South China area and Tethys area, combined with previously reported carbon and oxygen isotope values. Except for the extinction events at the end of the Permian, two significant changes were identified in brachiopod populations near the Cisuralian/Guadalupian and Guadalupian/Lopingian epoch boundaries in the Permian about 6⁃8 million years apart. The first change was evident in the mixed transitions of cold-water and warm-water species, and the latter is seen in various differentiation phenomena (individual enlargement, development of shell ornamentation and individual shrinkage, and shell thinning). The strengths of the ice age and monsoons were the main climatic factors, when temperature and nutrient sources jointly affected brachiopod diversity. Volcanism and plate movements changed the brachiopods’ ecological environment and caused regional differentiation. Sea-level changes were affected both by climate and crustal movement. In the study area, highly frequent sea-level changes constantly altered the living space of the brachiopods. The changes in shell shape and community structure succession of brachiopods were adaptations to different environments, showing the response of the biological environment to the paleoenvironment. -
Key words:
- brachiopods /
- macro evolution /
- community succession /
- diversity /
- Permian
-
图 2 中扬子新滩地区二叠纪腕足类种级延限
观测的数据:指实际产出的腕足类分异度;理想范围数据:指在实际产出的数据基础之上加上首现与末现层位出现的延限分子(如某一分子在一段剖面的上下都有产出,但中部并未产出)
Figure 2. The species⁃level range of Permian brachiopods in Xintan area, Middle Yangtze
(Observed data: actual observed data; Range⁃through data: ideally obtainable data)
图 3 中扬子区腕足类多样性曲线与华南腕足数据对比(I:Palaeofusulina sinensis带;II:Neoschwagerina haydeni带;III:Chusenella conicocylindrica带;IV:长兴组)
在N tot及N emsd中,均以实线及虚线来区分观测的数据及理想范围数据。N tot代表的是该时期内多样性的总和。N emsd与华南常驻属级多样性意义是相同的。E.吴家坪期为峨眉山火成岩喷发时期与吴家坪时期交界期
Figure 3. Comparison of brachiopod diversity curves in middle Yangtze region and brachiopod data in South China (Fusulines: I. Palaeofusulina sinensis. II. Neoschwagerina haydeni. III. Chusenella conicocylindrica. IV. Changhsingian Formation)
In Ntot and Nemsd, solid and dashed lines distinguish between observed and range⁃through data. Ntot represents the total diversity of the period. The E. Wuchaiapingian is an Emeishan volcanic event
-
[1] 陈洪德,王成善,刘文均,等. 华南二叠纪层序地层与盆地演化[J]. 沉积学报,1999,17(4):529-535. Chen Hongde, Wang Chengshan, Liu Wenjun, et al. 1999. Permian sequence stratigraphy and basin evolution in south of China[J]. Acta Sedimentologica Sinica, 1999, 17(4): 529-535. [2] 何卫红,唐婷婷,乐明亮,等. 华南南华纪一二叠纪沉积大地构造演化[J]. 地球科学:中国地质大学学报,2014,39(8):929-953. He Weihong, Tang Tingting, Yue Mingliang, et al. Sedimentary and tectonic evolution of Nanhuan-Permian in South China[J]. Earth Science:Journal of China University of Geosciences, 2014, 39(8): 928-953. [3] 宋海军,童金南. 二叠纪—三叠纪之交生物大灭绝与残存[J]. 地球科学,2016,41(6):901-918. Song Haijun, Tong Jinnan. Mass extinction and survival during the Permian-Triassic crisis[J]. Earth Science, 2016, 41(6): 901-918. [4] Shen S Z, Crowley J L, Wang Y, et al. Calibrating the end-Permian mass extinction[J]. Science, 2011, 334(6061): 1367-1372. [5] Black B A, Neely R R, Lamarque J F, et al. Systemic swings in end-Permian climate from Siberian traps carbon and sulfur outgassing[J]. Nature Geoscience, 2018, 11(12): 949-954. [6] Shen S Z, Henderson C M, Bowring S A, et al. High‐Resolution Lopingian (Late Permian) timescale of South China[J]. Geological Journal, 2010, 45(2/3): 122-134. [7] Shen S Z, Zhang H, Shi G R, et al. Early Permian (Cisuralian) global brachiopod palaeobiogeography[J]. Gondwana Research, 2013, 24(1): 104-124. [8] Burgess S D, Bowring S, Shen S. High-precision timeline for Earth’s most severe extinction[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(9): 3316-3321. [9] Henderson C M. Permian conodont biostratigraphy[J]. Geological Society, London, Special Publications, 2018, 450(1): 119-142. [10] Yuan D X, Shen S Z, Henderson C M, et al. Revised conodont-based integrated high-resolution timescale for the Changhsingian stage and End-Permian extinction interval at the Meishan Sections, South China[J]. Lithos, 2014, 204: 220-245. [11] Dimichele W A, Montañez I P, Poulsen C J, et al. Climate and vegetational regime shifts in the Late Paleozoic ice age earth[J]. Geobiology, 2009, 7(2): 200-226. [12] Yao X, Zhou Y Q, Hinnov L A. Astronomical forcing of a Middle Permian chert sequence in Chaohu, South China[J]. Earth and Planetary Science Letters, 2015, 422: 206-221. [13] Anderson R Y. Earth as diode: Monsoon source of the orbital~100 ka climate cycle[J]. Climate of the Past Discussions, 2010, 6(4): 1421-1452. [14] Jin Y G, Wang Y, Wang W, et al. Pattern of marine mass extinction near the Permian-Triassic boundary in South China[J]. Science, 2000, 289(5478): 432-436. [15] Song H, Tong J, Chen Z Q. Two episodes of foraminiferal extinction near the Permian-Triassic boundary at the Meishan section, South China[J]. Australian Journal of Earth Sciences, 2009, 56(6): 765-773. [16] Song H J, Wignall P B, Tong J N, et al. Two pulses of extinction during the Permian-Triassic crisis[J]. Nature Geoscience, 2013, 6(1): 52-56. [17] Wignall P B, Twitchett R J. Oceanic anoxia and the end Permian mass extinction[J]. Science, 1996, 272(5265): 1155-1158. [18] Song H J, Wignall P B, Tong J N, et al. Geochemical evidence from bio-apatite for multiple oceanic anoxic events during Permian-Triassic transition and the link with end-Permian extinction and recovery[J]. Earth and Planetary Science Letters, 2012, 353-354: 12-21. [19] Song H J, Wignall P B, Chu D L, et al. Anoxia/high temperature double whammy during the Permian-Triassic marine crisis and its aftermath[J]. Scientific Reports, 2015, 4: 4132. [20] Algeo T J, Chen Z Q, Fraiser M L, et al. Terrestrial-marine teleconnections in the collapse and rebuilding of Early Triassic marine ecosystems[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2011, 308(1/2): 1-11. [21] Song H J, Wignall P B, Tong J N, et al. Integrated Sr isotope variations and global environmental changes through the Late Permian to Early Late Triassic[J]. Earth and Planetary Science Letters, 2015, 424: 140-147. [22] Joachimski M M, Lai X L, Shen S Z, et al. Climate warming in the Latest Permian and the Permian-Triassic mass extinction[J]. Geology, 2012, 40(3): 195-198. [23] Shen S Z, Zhang H, Li W Z, et al. Brachiopod diversity patterns from Carboniferous to Triassic in South China[J]. Geological Journal, 2006, 41(3/4): 345-361. [24] Golonka J, Ford D. Pangean (Late Carboniferous-Middle Jurassic) paleoenvironment and lithofacies[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2000, 161(1/2): 1-34. [25] Xu Y G, Wei X, Luo Z Y, et al. The Early Permian Tarim Large Igneous province: Main characteristics and a plume incubation model[J]. Lithos, 2014, 204: 20-35. [26] Zhou M F, Malpas J, Song X Y, et al. A temporal link between the Emeishan large igneous province (SW China) and the end-Guadalupian mass extinction[J]. Earth and Planetary Science Letters, 2002, 196(3/4): 113-122. [27] Shellnutt J G. The Emeishan large igneous province: A synthesis[J]. Geoscience Frontiers, 2014, 5(3): 369-394. [28] 殷鸿福,黄思骥,张克信,等. 华南二叠纪—三叠纪之交的火山活动及其对生物绝灭的影响[J]. 地质学报,1989,63(2):169-180. Yin Hongfu, Huang Siji, Zhang Kexin, et al. Volcanism at the Permian-Triassic boundary in South China and its effects on mass extinction[J]. Acta Geologica Sinica, 1989, 63(2): 169-180. [29] Gao Q L, Zhang N, Xia W C, et al. Origin of volcanic ash beds across the Permian⁃Triassic boundary, Daxiakou, South China: Petrology and U⁃Pb age, trace elements and Hf-isotope composition of zircon[J]. Chemical Geology, 2013, 360-361: 41-53. [30] Reichow M K, Pringle M S, Al'Mukhamedov A I, et al. The timing and extent of the eruption of the Siberian Traps large igneous province: Implications for the end-Permian environmental crisis[J]. Earth and Planetary Science Letters, 2009, 277(1/2): 9-20. [31] Burgess S D, Bowring S A. High-precision geochronology confirms voluminous magmatism before, during, and after Earth’s most severe extinction[J]. Science Advances, 2015, 1(7): e1500470. [32] Zhang H, Cao C Q, Liu X L, et al. The terrestrial end-Permian mass extinction in South China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2016, 448: 108-124. [33] 王成善,陈洪德,寿建峰,等. 中国南方二叠纪层序地层划分与对比[J]. 沉积学报,1999,17(4):499-509. Wang Chengshan, Chen Hongde, Shou Jianfeng, et al. Characteristics and correlation of Permian depositional sequences in South China[J]. Acta Sedimentologica Sinica, 1999, 17(4): 499-509. [34] Lehrmann D J, Enos P, Payne J L, et al. Permian and Triassic depositional history of the Yangtze platform and great bank of Guizhou in the Nanpanjiang Basin of Guizhou and Guangxi, South China[J]. Albertiana, 2005, 33: 147-166. [35] 冯少南,许寿永,林甲兴,等. 长江三峡地区生物地层学(3)晚古生代分册[M]. 北京:地质出版社,1984,63-109. Feng Shaonan, Xu Shouyong, Lin Jiaxing, et al. Biostratigraphy in the Three Gorges of the Yangtze River (3) Late Paleozoic[M]. Beijing: Geological Publishing House, 1984, 63-109. [36] Rong J Y, Shen S Z, Zhan R B, et al. Phanerozoic Brachiopod Genera of China, Volume 2[M]. Beijing, Beijing: Science Press, 2017:1-38. [37] Shen S Z, Zhang H, Zhang Y C, et al. Permian integrative stratigraphy and timescale of China[J]. Science China Earth Sciences, 2019, 62(1): 154-188. [38] 陈清,樊隽轩,Melchin M J. 古生物多样性统计方法及其适用性分析[J]. 古生物学报,2012,51(4):445-462. Chen Qing, Fan Junxuan, Melchin M J. Methods for paleobiodiversity measurement and case studies of their applicability[J]. Acta Palaeontologica Sinica, 2012, 51(4): 445-462. [39] 尚庆华,金玉玕. 二叠纪腕足动物地理区系演化特征[J]. 古生物学报,1997,36(1):93-121. Shang Qinghua, Jin Yugan. Quantitative evaluation on paleozoogeographic evidences of Permian brachiopods[J]. Acta Palaeontologica Sinica, 1997, 36(1): 93-121. [40] Archbold N W, Pigram C J, Ratman N, et al. Indonesian Permian brachiopod fauna and Gondwana⁃South-East Asia relationships[J]. Nature, 1982, 296(5857): 556-558. [41] Huang X, Aretz M, Zhang X H, et al. Pennsylvanian-early Permian palaeokarst development on the Yangtze Platform, South China, and implications for the regional sea-level history[J]. Geological Journal, 2018, 53(4): 1241-1262. [42] Haq B U, Schutter S R. A chronology of Paleozoic sea-level changes[J]. Science, 2008, 322(5898): 64-68. [43] Chen B, Joachimski M M, Shen S Z, et al. Permian ice volume and palaeoclimate history: Oxygen isotope proxies revisited[J]. Gondwana Research, 2013, 24(1): 77-89. [44] Bagherpour B, Bucher H, Schneebeli-Hermann E, et al. Early Late Permian coupled carbon and strontium isotope chemostratigraphy from South China: Extended Emeishan volcanism?[J]. Gondwana Research, 2018, 58: 58-70. [45] Korte C, Jasper T, Kozur H W, et al. δ18O and δ13C of Permian brachiopods: A record of seawater evolution and continental glaciation[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2005, 224(4): 333-351. [46] Rau G H, Takahashi T, Des Marais D J, et al. Latitudinal variations in plankton δ13C: Implications for CO2 and productivity in past oceans[J]. Nature, 1989, 341(6242): 516-518. [47] Meyer K M, Yu M, Jost A B, et al. δ13C evidence that high primary productivity delayed recovery from end-Permian mass extinction[J]. Earth and Planetary Science Letters, 2011, 302(3/4): 378-384. [48] Westerhold T, Bickert T, Röhl U. Middle to Late Miocene oxygen isotope stratigraphy of ODP site 1085 (SE Atlantic): New constrains on Miocene climate variability and sea-level fluctuations[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2005, 217(3/4): 205-222. [49] Beauchamp B, Baud A. Growth and demise of Permian biogenic chert along northwest Pangea: Evidence for end-Permian collapse of thermohaline circulation[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2002, 184(1/2): 37-63. [50] Belasky P, Stevens C H, Hanger R A. Early Permian location of western North American terranes based on brachiopod, fusulinid, and coral biogeography[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2002, 179(3/4): 245-266. [51] Zuschin M, Mayrhofer S. Brachiopods from cryptic coral reef habitats in the northern Red Sea[J]. Facies, 2009, 55(3): 335-344. [52] Shields C A, Kiehl J T. Monsoonal precipitation in the Paleo-Tethys warm pool during the Latest Permian[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2018, 491: 123-136. [53] 殷鸿福,喻建新,罗根明,等. 地史时期生物对冰室气候形成的作用[J]. 地球科学,2018,43(11):3809-3822. Yin Hongfu, Yu Jianxin, Luo Genming, et al. Biotic influence on the formation of icehouse climates in geologic history[J]. Earth Science, 2018, 43(11): 3809-3822. [54] 魏信祥,张雄华,黄兴,等. 吐哈盆地中二叠世古气候重建:来自木化石年轮的证据[J]. 地球科学,2016,41(10):1771-1780. Wei Xinxiang, Zhang Xionghua, Huang Xing. et al. Palaeoclimate reconstruction of Middle Permian in Tuha Basin: Evidence from the fossil wood growth rings[J]. Earth Science, 2016, 41(10): 1771-1780. [55] 刘荞菲. 瘤状灰岩的分类及成因研究[J]. 四川有色金属,2015,(2):26-28,39. Liu Qiaofei. The classification and the causes of nodular limestone[J]. Sichuan Nonferrous Metals, 2015, (2): 26-28, 39. [56] 詹仁斌,靳吉锁,刘建波. 奥陶纪生物大辐射研究:回顾与展望[J]. 科学通报,2013,58(33):3357-3371. Zhan Renbin, Jin Jisuo, Liu Jianbo. Investigation on the great Ordovician biodiversification event (GOBE): Review and prospect[J]. Chinese Science Bulletin, 2013, 58(33): 3357-3371. [57] 卜建军,何卫红,张克信,等. 古亚洲洋的演化:来自古生物地层学方面的证据[J]. 地球科学,2020,45(3):711-727. Bu Jianjun, He Weihong, Zhang Kexin, et al. Evolution of the Paleo-Asian Ocean: Evidences from paleontology and stratigraphy[J]. Earth Science, 2020, 45(3): 711-727. [58] Fang Q, Wu H C, Hinnov L A, et al. Astronomical cycles of Middle Permian Maokou Formation in South China and their implications for sequence stratigraphy and paleoclimate[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 474: 130-139. [59] Isozaki Y, Aljinović D, Kawahata H. The Guadalupian (Permian) Kamura event in European Tethys[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2011, 308(1/2): 12-21. [60] 龚志军,谢平,阎云君. 底栖动物次级生产力研究的理论与方法[J]. 湖泊科学,2001,13(1):79-88. Gong Zhijun, Xie Ping, Yan Yunjun. Theories and methods of studies on the secondary production of zoobenthos[J]. Journal of Lake Sciences, 2001, 13(1): 79-88. [61] 廖卓庭. 贵州西部上二叠统腕足化石[M]//中国科学院南京地质古生物研究所. 黔西滇东晚二叠世含煤地层和古生物群. 北京:科学出版社,1980:241-277. [ Liao Zhuoting. Fossil of Upper Permian brachiopod in western Guizhou[M]//Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences. Late Permian coal-bearing strata and paleobiota in eastern Guizhou, Yunnan and eastern Yunnan. Beijing: Science Press, 1980: 241-277.] [62] 廖卓庭. 中国南部长兴阶的腕足动物组合带及二叠、三叠纪混生动物群中的腕足动物[J]. 地层学杂志,1979,3(3):200-207. Liao Zhuoting. Brachiopod combination zone of Changxingian in southern China and brachiopods in mixed Permian and Triassic faunas[J]. Acta Stratigraphy Sinica, 1979, 3(3): 200-207. [63] Wang X D, Sugiyama T. Diversity and extinction patterns of Permian coral faunas of China[J]. Lethaia, 2000, 33(4): 285-294. [64] 张阳,何卫红. 华南二叠纪腕足长身贝类的形态演化规律[J]. 中国科学(D辑):地球科学,2008,38(12):1521-1532. Zhang Yang, He Weihong. The morphological evolution of the Permian brachiopod long shellfish in South China[J]. Science China (Seri. D): Earth Sciences, 2008, 38(12): 1521-1532. [65] Clapham M E, Shen S Z, Bottjer D J, et al. The double mass extinction revisited: Reassessing the severity, selectivity, and causes of the end-Guadalupian biotic crisis (Late Permian)[J]. Paleobiology, 2009, 35(1): 32-50. [66] 韦雪梅,韦恒叶,邱振. 瓜德鲁普末期生物灭绝是由峨眉山大火成岩省(Lip)引起的吗?[J]. 沉积学报,2016,34(3):436-451. Wei Xuemei, Wei Hengye, Qiu Zhen. Was the End-Guadalupian mass extinction caused by the Emeishan LIP eruption?[J]. Acta Sedimentologica Sinica, 2016, 34(3): 436-451. [67] Clapham M E, Bottjer D J. Permian marine paleoecology and its implications for large-scale decoupling of brachiopod and bivalve abundance and diversity during the Lopingian (Late Permian)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2007, 249(3/4): 283-301. [68] Weidlich O, Kiessling W, Flügel E. Permian-Triassic boundary interval as a model for forcing marine ecosystem collapse by long-term atmospheric oxygen drop[J]. Geology, 2003, 31(11): 961-964. [69] Clapham M E, Payne J L. Acidification, anoxia, and extinction: A multiple logistic regression analysis of extinction selectivity during the Middle and Late Permian[J]. Geology, 2011, 39(11): 1059-1062. [70] He W H, Shi G R, Twitchett R J, et al. Late Permian marine ecosystem collapse began in deeper waters: Evidence from brachiopod diversity and body size changes[J]. Geobiology, 2015, 13(2): 123-138. [71] Montenegro A, Spence P, Meissner K J, et al. Climate simulations of the Permian‐Triassic boundary: Ocean acidification and the extinction event[J]. Paleoceanography, 2011, 26(3): PA3207. [72] Zhang K X, Tong J N, Yin H F, et al. Sequence stratigraphy of the Permian–Triassia boundary section of Changxing, Zhejiang, southern China[J]. Acta Geologica Sinica, 1997, 71(1): 90-103. [73] Kawamura T, Machiyama H. A Late Permian coral reef complex, South Kitakami Terrane, Japan[J]. Sedimentary Geology, 1995, 99(3/4): 135-150. [74] Molinos J G, Burrows M T, Poloczanska E S. Ocean currents modify the coupling between climate change and biogeographical shifts[J]. Scientific Reports, 2017, 7(1): 1332. [75] Pringle J M, Byers J E, He R Y, et al. Ocean currents and competitive strength interact to cluster benthic species range boundaries in the coastal ocean[J]. Marine Ecology Progress Series, 2017, 567: 29-40. [76] 沈树忠,张华. 什么引起五次生物大灭绝?[J]. 科学通报,2017,62(11):1119-1135. Shen Shuzhong, Zhang Hua. What caused the five mass extinctions?[J]. Chinese Science Bulletin, 2017, 62(11): 1119-1135. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 758
- HTML全文浏览量: 168
- PDF下载量: 66
- 被引次数: 0


