
HTML
-
近20年来对现代植物叶片的碳同位素变化及其控制因素的研究取得了一系列重要的进展,其中有两个方面的研究成果引起地学界古气候研究领域专家和学者的普遍关注。一是Kohn[1]和Diefendorf et al.[2]发现陆生C3型植物的叶片碳同位素(δ13C叶片)受平均年降水量控制,Stein et al.[3]研究证实现代土壤有机质碳同位素(δ13C土壤)与平均年降水量之间也呈负相关关系。而Schubert et al.[4]和Cui et al.[5]提出陆生C3型植物的δ13C叶片记录了大气pCO2浓度变化。如果采用实验室分离和测试方法,提取沉积有机质中保存的陆地植物叶片的δ13C叶片信号,这时引用现代植物的δ13C叶片与降雨量和大气pCO2浓度的数学函数关系式,则可以定量重建古气候的干湿变化历史,描述驱动冰室和温室气候状态转换过程中古大气CO2浓度的变化过程。解决这一问题有两种手段,一是直接测定地层剖面中植物化石叶片的碳同位素δ13C叶片[6⁃9],二是测定沉积有机质中长链正构烷烃分子单体的碳同位素,因为高碳数长链正构烷烃nC27、nC29和nC31来源于陆地生长的木本和草本植物,这些长链正构烷烃单体分子的碳同位素平均值标记为δ13C nC27-nC29-nC31,保存了古植物叶片的碳同位素δ13C叶片信号[10⁃14]。目前的问题是,古植物叶片一般在地层剖面上分散出现,它记录的是沉积期一个短时段的气候状态,而长链高碳数正构烷烃分子的碳同位素在富有机质地层剖面可以连续采样,且正构烷烃分子δ13C正构烷烃较植物叶片δ13C叶片偏负。据Tipple et al.[12]的资料,现代C3植物叶片δ13C平均值-27.8‰,正构烷烃δ13C nC29、δ13C nC31平均值分别为-33.3‰、-33.9‰,正构烷烃碳同位素比现代植物叶片碳同位素偏负-5.5‰~-6.1‰;另外,Liu et al.[15]统计发现,全球现代陆生植物正构烷烃δ13C nC27、δ13C nC29、δ13C nC31平均值分别-34.1‰、-35.0‰,-35.3‰,δ13C正构烷烃较现代C3植物叶片δ13C平均值-27.8‰偏负达-6.3‰~-7.5‰。因此,如何选择参数将δ13C正构烷烃校对为δ13C叶片值面临挑战。目前,关于古植物化石与对应的同一件化石样品围岩的可溶有机质正构烷烃碳同位素之间关系的研究资料极为稀少,缺乏实测植物化石碳同位素与含化石岩层全岩有机质(δ13CTOC)或干酪根(δ13C干酪根)以及δ13C nC27-nC29-nC31的分析对比资料。本文选择植物化石丰富的地层剖面,同时采集植物叶片、茎秆化石和含植物化石岩层的样品,室内同步分析古植物化石和岩石中可溶有机质中正构烷烃的碳同位素值,标定δ13C古植物与δ13C nC27-nC29-nC31的校对值。同时,为了对比δ13CTOC或δ13C干酪根与高碳数长链正构烷烃δ13C nC27-nC29-nC31之间的差值,选择柴达木盆地英雄岭地区狮68井下干柴沟组盐湖相开展了δ13C干酪根与可溶有机质氯仿沥青(δ13C氯仿沥青)、饱和烃(δ13C饱和烃)以及正构烷烃单体分子碳同位素的研究,以期为重建地质历史时期地层剖面δ13C古植物演化过程提供资料。
-
植物化石采集点为柴达木盆地东北缘大煤沟地区,侏罗纪地层划分为下侏罗统小煤沟组、中侏罗统大煤沟组、采石岭组,上侏罗统红水沟组。这一地区是中国侏罗纪产植物化石最丰富的剖面之一,青海省地质科学研究所和中国科学院南京地质古生物研究所李佩娟等[16]曾出版有关侏罗纪植物群专著,描述有植物化石63属198种。新生代植物化石采自宋博文[17]报道的大红沟剖面,侏罗纪化石采自大煤沟组煤层顶底板和夹层(图1)。野外采集的植物化石经南京地质调查中心钱迈平研究员鉴定,侏罗纪地层见有费尔干似银杏(Ginkgoites ferganensis)、奇丽楔拜拉(Spenobaiera spectabilis)、布列亚锥叶蕨(Coniopteris burejensis)、海庞枝脉蕨(Cladophlebis haiburnensis)、海西州新芦木(Neocalamites haixizhouensis)、乌兰枝脉蕨(Cladophlebis ulanensis)、美丽锥叶蕨(Coniopteris spectabilis)、中华蒂尔姆叶(Tyrmia sinensis)、青海假托勒利叶(Pseudotorellia qinghaiensis),古近纪地层见有豆科植物(Leguminosae)和木贼(Equisetum sp.)化石碎片。

Figure 1. Location of sampling sites in the Qaidam Basin
-
样品分析和测试分别在国家地质实验测试中心和华北油田勘探开发研究院油田实验室开展。野外采集的植物化石,室内首先用蒸馏水清洗化石表面的杂质,放入通风橱中自然晾干,然后用刻刀刮削植物实体化石,并采集粉末。粉末样品经粉碎、过筛,同时对盛装样品的坩埚进行高温煅烧,在1 000 ℃条件下灼烧2 h以上去除杂物。称取约0.1 g粉末样品置于高温煅烧冷却后的坩埚中,在60 ℃环境下缓慢滴入配制好的稀盐酸溶液(盐酸∶水=1∶7,体积比),化石样品与盐酸充分反应直至无气泡产生。最后用蒸馏水反复淋洗坩埚,直至滤液为中性,洗样完毕后置于60 ℃烘箱内干燥以备开展碳同位素测定。
化石围岩样品首先剥离风化面,粉碎、过筛处理至100~200 目,预处理方式参照植物化石处理流程。植物化石和围岩样品碳同位素测定采用EA-IRMS联用系统测量,同位素比质谱仪型号为Elementar Isoprime Precision。在样品分析前开展标准样品测试,以确保仪器处于良好工作状态。每测量十个样品,至少进行一次平行样品及标准样品的测试,以保证仪器运行稳定性,标样分别为GBWE04407、GBWE04408和USGS24。
碳同位素组成测试流程:在锡杯中称取适量处理后的样品,放入自动进样器,启动元素分析仪和同位素质谱仪的采集程序,样品经元素分析仪充分燃烧后,生成CO2并随载气流进入同位素质谱仪进行碳同位素组成测定,碳同位素重复测试的误差控制在±0.3‰以内。设定元素分析仪燃烧管温度设定为950 ℃,还原管温度设定为600 ℃。狮68井干酪根样品制备按标准SY/T 5123—1995执行。制备流程如下:蒸馏水浸泡、盐酸或氢氟酸酸处理、碱处理、黄铁矿处理、重液浮选、冷冻干燥,氯仿清洗可溶有机质和烧失量的测定。经多步反应后,制备后的干酪根样品烧失量大于75%,存放于60 ℃烘箱中干燥,最后进行干酪根碳同位素分析。δ13CTOC和δ13C干酪根实验测定流程和误差详见参考文献[18]。
植物化石围岩的氯仿沥青分离采用索氏抽提方法:首先将粉末样品约50 g装入滤纸袋中,通入氯仿溶剂对岩石粉末中的有机质进行萃取(标准SY/T 5118—1995),第二步对氯仿沥青进行族组分分离(标准SY/T 5119—1995),最后对饱和烃馏分进行气相色谱分析(标准SY/T 5258—1995)。色谱分析采用Agilent 6890 N型气相色谱仪,色谱柱为HP-5石英弹性毛细管色谱柱(30 m×0.32 mm×0.25 μm),载气为氦气。色谱柱初温60 ℃恒温5 min,升温速度为3 ℃/min,终温290 ℃恒温20 min。
单体正构烷烃碳同位素分析采用Delta V Advantage同位素质谱仪,实验方法及流程按照《质谱分析方法通则》(GB/T 6041—2002)执行。色谱仪柱箱中的色谱柱为DB-5MS硅熔融毛细管色谱柱,载气为N2,无分流进样为1~1.2 μL,升温程序为:色谱仪柱箱初始温度为60 ℃,以10 ℃/min升至250 ℃,后以6 ℃/min升至320 ℃,恒温7 min;质谱仪初始温度为70 ℃,以4 ℃/min持续升至320 ℃,持续10 min。正构烷烃单体碳同位素计算公式为δ13C=[(R样品/R标样)-1]×1 000。13C/12C采用VPDB标准的千分含量,分析误差小于±0.2‰。表1和表2分别是侏罗纪大煤沟和古近纪大红沟地区以及狮子沟地区狮68井钻井剖面沉积有机质实验分析和统计数据表。
序号 样品编号 样品类型 属种名称 植物化石样品碳同位素分析值/‰ 化石围岩样品正构烷烃分子化合物碳同位素值/‰ 植物化石 化石围岩 氯仿沥青 饱和烃 nC23 nC25 nC27 nC29 nC31 nC33 1 DP93f 叶片化石 费尔干似银杏 -23.0 -22.2 -27.1 -27.1 -26.7 -27.8 -28.0 -27.9 -27.1 -24.7 2 DP31f 叶片化石 奇丽楔拜拉 -23.0 -22.6 -25.6 -26.7 -27.3 -27.8 -27.9 -28.2 -29.1 -26.7 3 DP67f 叶片化石 奇丽楔拜拉 -23.0 -22.2 -27.1 -27.1 -26.8 -27.8 -28.0 -27.9 -28.2 -26.6 4 DP22f 叶片化石 奇丽楔拜拉 -22.8 -22.7 -27.4 -27.0 -26.6 -26.9 -27.4 -27.7 -27.5 -25.0 5 DP50f 叶片化石 海西州新芦木 -23.6 -24.4 -25.5 -27.1 -26.6 -27.3 -27.9 -28.4 -26.8 6 DP123 叶片化石 海西州新芦木 -25.5 -23.5 -29.3 -28.2 -29.7 -28.6 -28.6 -28.8 -28.6 7 DMG29 叶片化石 布列亚锥叶蕨 -23.2 -23.1 -26.2 -27.4 -27.3 -27.6 -28.3 -28.9 -28.4 -26.3 8 DP90 叶片化石 美丽锥叶蕨 -23.0 -23.5 -25.8 -26.6 -26.7 -27.4 -28.0 -28.9 -29.1 -26.5 9 DP106f 叶片化石 海庞枝脉蕨 -22.7 -22.8 -29.2 -29.0 -28.5 -28.7 -29.7 -30.5 -29.9 10 DP112f 叶片化石 乌兰枝脉蕨 -23.2 -23.6 -25.6 -27.4 -27.2 -27.7 -28.2 -30.6 -28.9 11 SBWf 叶片化石 豆科植物 -26.2 -27.1 -31.0 -29.4 -30.0 -30.4 -31.2 -31.7 -27.6 -33.1 12 SH6f 茎秆化石 木贼植物 -23.8 -23.5 -28.3 -27.1 -29.5 -28.1 -28.2 -29.9 -28.5 -30.1 13 SYQ2f 茎秆化石 木贼植物 -23.8 -23.6 -29.5 -27.1 -29.8 -28.6 -28.5 -28.9 -28.9 -31.2 14 SYQ3f 茎秆化石 木贼植物 -24.7 -23.5 -29.6 -27.2 -30.0 -28.5 -28.6 -29.4 -29.5 -30.5 注: 编号DP和DMG样品采自大煤沟侏罗纪地层分布区,编号SBW、SH和SYQ样品采自大红沟古近纪地层分布区。Table 1. Carbon isotopes analysis data of sedimentary organic matter from plant fossils and sampling rocks of the Jurassic Dameigou Formation and Paleogene Shangganchaigou Formation in the Qaidam Basin
统计值/类别 δ13C干酪根 δ13C氯仿沥青 δ13C饱和烃 正构烷烃 短链正构烷烃 长链正构烷烃 平均值 平均值 平均值 δ13C nC14~nC32 δ13C nC14~nC21 δ13C nC22~nC32 最小值 -28.4 -27.7 -28.8 -31.3 -30.6 -32.9 平均值 -25.8 -25.9 -27.6 -29.6 -29.0 -30.1 最大值 -23.2 -24.0 -25.9 -27.2 -27.0 -27.3 统计值/类别 短链奇偶碳数 短链奇数碳 中链奇偶碳数 中链奇碳数 长链奇偶碳数 长链奇碳数 正构烷烃 正构烷烃 正构烷烃 正构烷烃 正构烷烃 正构烷烃 δ13C nC14~nC20 δ13C nC15-nC17-nC19 δ13C nC21~nC25 δ13C nC21-nC23-nC25 δ13C nC26~nC32 δ13C nC27-nC29-nC31 最小值 -30.4 -30.5 -32.1 -32.1 -33.7 -34.3 平均值 -28.9 -29.1 -29.7 -29.7 -30.4 -30.9 最大值 -27.2 -27.4 -26.2 -26.2 -27.7 -28.7 Table 2. Carbon isotopes analysis data of sedimentary organic matter from the Paleogene Xiaganchaigou Formation from well Shi 68 in the Qaidam Basin
-
野外采集的14件植物叶片和茎秆化石样品碳同位素(δ13C古植物)分布范围为-26.2‰~-22.7‰,平均值为-23.7‰。侏罗纪大煤沟组10件植物叶片样品(编号DP和DMG)碳同位素平均值为-23.3‰,古近纪下干柴沟组4件植物叶片和茎秆样品(编号SBW、SH、SYQ)碳同位素平均值为-24.6‰。据Richey et al.[19]等统计资料,新生代和中生代植物叶片化石δ13C古植物平均值分别为-24.4‰和-25.4‰,植物茎秆化石δ13C古植物平均值分别为-25.3‰和-24.0‰,研究区侏罗纪和古近纪δ13C古植物平均值较文献报道的中生代和新生代古植物化石δ13C古植物平均值-24.4‰和-25.9‰偏正+0.7‰~+2.2‰。
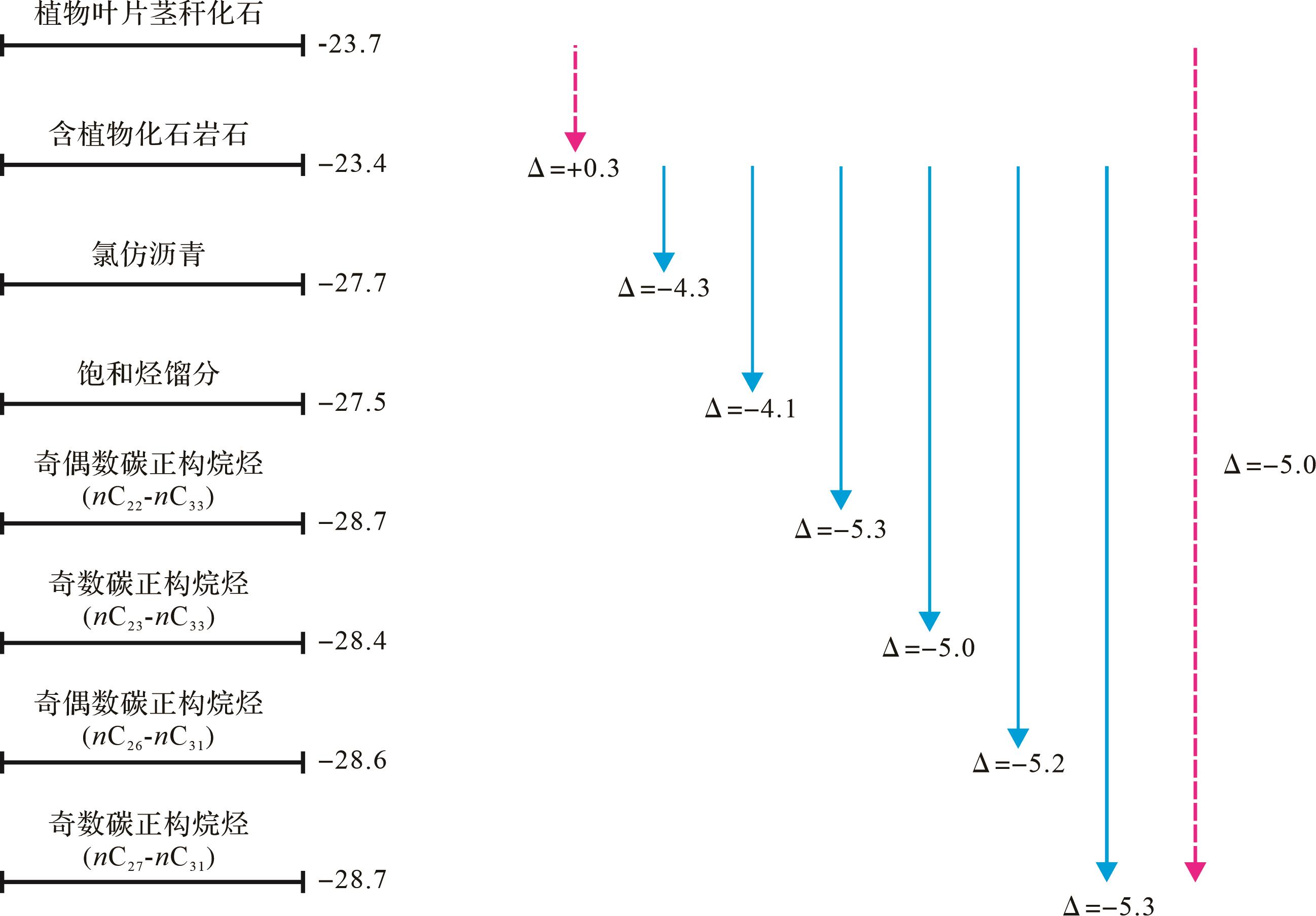
观察和对比δ13C古植物和δ13CTOC以及岩石中分离的δ13C氯仿沥青、δ13C饱和烃,可以发现δ13CTOC较δ13C古植物偏正+0.3‰,而氯仿沥青和饱和烃组分碳同位素较δ13CTOC偏负-4.3‰和-4.1‰(图2)。

Figure 2. Average difference value comparison of carbon isotopes of individual n⁃alkanes from plant fossils and rock samplesof the Jurassic Dameigou Formation and Paleogene Shangganchaigou Formation in the Qaidam Basin
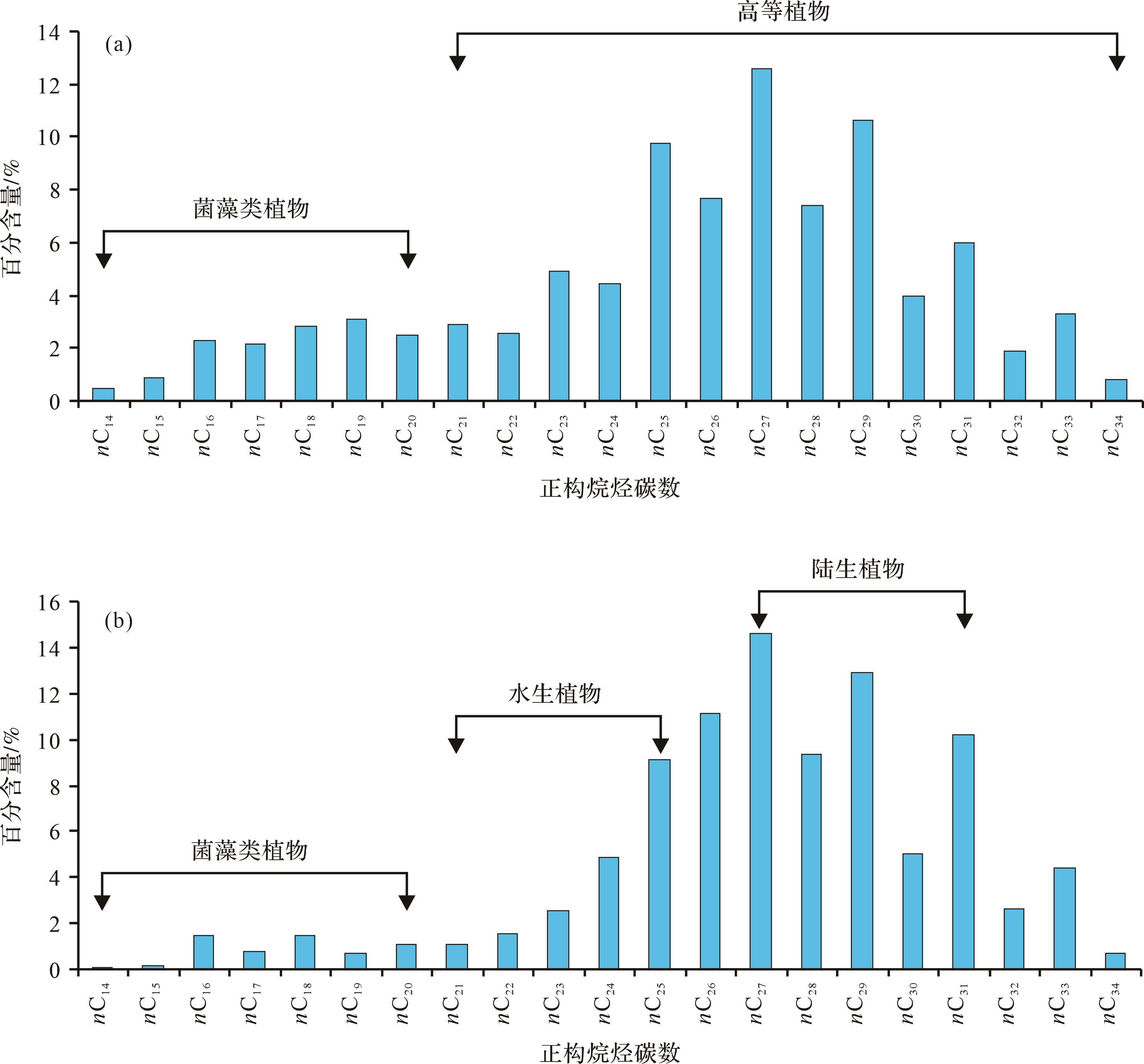
采用索氏抽提法从古植物化石的岩石样品中抽提的可溶有机质氯仿沥青,进一步通过柱层析法分离的饱和烃组分的色谱分析结果如图3所示。分析侏罗纪大煤沟组10件样品和古近纪上干柴沟组4件样品的饱和烃色谱图可以发现,正构烷烃碳数分布区间介于nC14~nC34,低碳数短链正构烷烃(nC14~nC20)含量低,且奇碳数和偶碳数正构烷烃丰度变化不明显,而高碳数长链正构烷烃(nC21~nC34)含量高,出现奇碳数大于偶碳数正构烷烃含量的分布特点,奇偶优势指标OEP变化范围为0.89~2.37,碳优势参数CPI波动介于1.11~2.68。色谱图中主峰碳数为nC27,其次为nC29和nC25,nC31和nC33丰度最小。一般认为,短链奇数碳正构烷烃nC15和nC17来源于水生菌藻类植物,nC23和nC25是水生高等植物的标志,而高碳数长链正构烷烃nC27、nC29和nC31主峰碳是陆生高等植物的特征,nC27和nC29起源于木本植物,nC31在草本植物中含量最高[20⁃24]。

Figure 3. Gas chromatograms of saturated hydrocarbon fraction from extractable organic matter from plant leaf and stalk fossil rock samples of the Jurassic Dameigou Formation and Paleogene Shangganchaigou Formation in the Qaidam Basin
植物化石围岩中δ13C正构烷烃分布范围和平均值(表1、图2)显示正构烷烃单体分子中记录的高等植物(水生植物和陆生植物)奇碳数和偶碳数正构烷烃δ13C nC26-nC31平均值为-28.6‰,奇碳数正构烷烃δ13C nC23-nC33平均值为-28.4‰,分别较δ13CTOC偏负-5.2‰和-5.0‰。陆生高等植物特征标志的δ13C nC27-nC29-nC31为-28.7‰,较δ13C古植物偏负-5.0‰。从植物化石产出地层来看,侏罗纪植物化石有机质碳同位素较含化石岩石δ13C nC27-nC29-nC31平均值偏正+5.1‰,古近纪植物化石有机质碳同位素较含化石岩石奇碳数正构烷烃碳同位素δ13C nC27-nC29-nC31平均值偏正+4.7‰。
-
现代盐湖沉积的有机质,主要来源于湖盆周缘高地的陆生植物输入,而很少有水生藻类植物有机质的出现,中国西北地区茶卡盐湖和察尔汗盐湖就是这类高盐度湖泊的典型实例。据郭金春等[24]和段毅等[25]研究,现代盐湖底部沉积物中正构烷烃分布与湖盆周缘陆地木本和草本植物一致,显示以nC25、nC27、nC29和nC31为主峰的长链高碳数正构烷烃分布为优势。然而,在与蒸发岩共生的海相和陆相沉积地层中,菌藻类成因的正构烷烃占有很大的比例,而且出现偶碳数正构烷烃含量大于奇碳数正构烷烃的分布形式,并以nC22高峰为标志[26⁃32]。这类高盐度湖泊环境、菌藻类和陆生植物混合来源以及经历复杂埋藏成岩过程的正构烷烃分子碳同位素是否能够记录陆生古植物的碳同位素信号仍不确定,我们选择柴达木盆地西部狮68井岩屑样品进行验证。在狮68井下干柴沟组上段(E

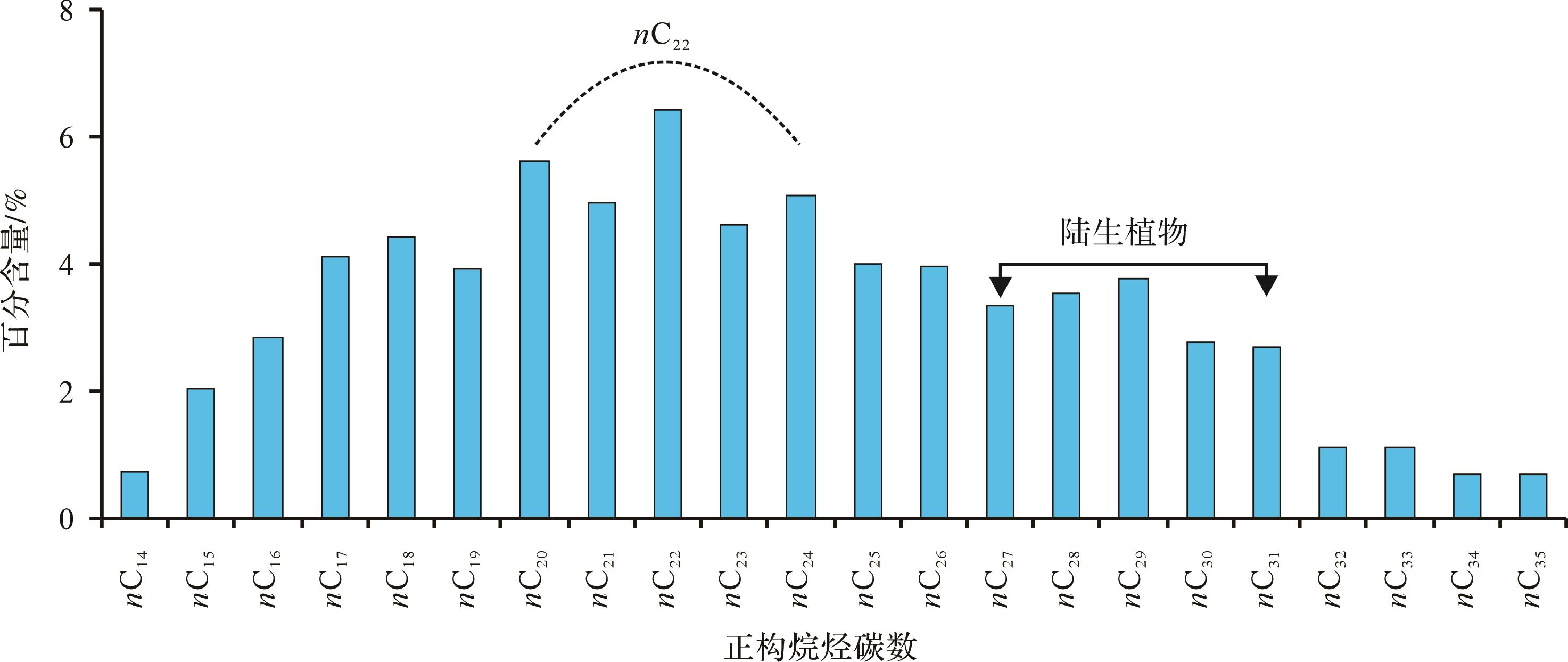
Figure 4. Gas chromatograms of saturated hydrocarbon fraction from black mudstones interlayered with alternative evaporitic deposits from the saline lacustrine depositional environment of well Shi 68
可以发现,正构烷烃碳数分布介于nC14~nC35,与侏罗纪大煤沟组有所不同,这些样品的正构烷烃主峰碳以偶数碳nC22为标志,次主峰有nC20和nC24,低碳数正构烷烃nC14~nC19和高碳数正构烷烃nC25~nC35部分奇偶优势不明显(图4)。奇偶优势指标OEP变化范围为0.70~1.41,碳优势参数CPI波动区间为0.37~0.75。偶碳数正构烷烃异常的出现一般认为与细菌微生物有关,它是高盐度海湾和盐湖蒸发岩地层沉积有机质正构烷烃分布普遍存在的特征[33⁃36]。
狮68井δ13C干酪根分布范围为-28.4‰~-23.2‰,平均值为-25.8‰(图5)。地表大红沟剖面下干柴沟组δ13C古植物平均值为-24.6‰,较干酪根碳同位素偏正+1.2‰,而狮68井的δ13C干酪根平均值与Richey et al.[18]报道的新生代煤层和木化石碳同位素-25.4‰、-25.3‰相近。岩屑样品δ13C氯仿沥青和δ13C饱和烃平均值分别为-25.9‰和-27.6‰,二者较δ13C干酪根平均值偏负-0.1‰和-1.8‰。对比δ13C干酪根、δ13C氯仿沥青、δ13C饱和烃变化区间发现,δ13C氯仿沥青和δ13C饱和烃分布区间(即最大值至最小值)较δ13C干酪根小。

Figure 5. Difference values comparison of carbon isotopes of separated kerogen and individual n⁃alkanes from extractable organic matter of the Xiaganchaigou Formation black mudstones of well Shi 68
狮68井高等植物包括水生植物和陆生植物奇偶数正构烷烃单体分子碳同位素(δ13C nC22-nC32)平均值为-30.1‰,奇数碳正构烷烃单体分子碳同位素(δ13C nC23-nC31)平均值为-30.5‰,二者较δ13C干酪根偏负-4.3‰和-4.7‰。水生植物常见的nC23和nC25正构烷烃单体分子平均值为-30.0‰,没有发现因现代湖泊水体中沉水植物存在而出现正的碳同位素异常[37⁃42]。陆生植物特征长链奇偶碳数正构烷烃单体分子碳同位素(δ13C nC26-nC32)平均值为-30.4‰,δ13C nC27-nC29-nC31平均值为-30.9‰。因为古植物叶片和茎秆之间碳同位素存在±1.1‰~±1.4‰变化[19,42],全岩有机质δ13CTOC较化石叶片δ13C化石叶片偏正0.3‰,为了简化计算,可采用5.0‰作为δ13C nC27-nC29-nC31的校对值,即δ13C古植物=δ13C nC27-nC29-nC31+5.0‰。
-
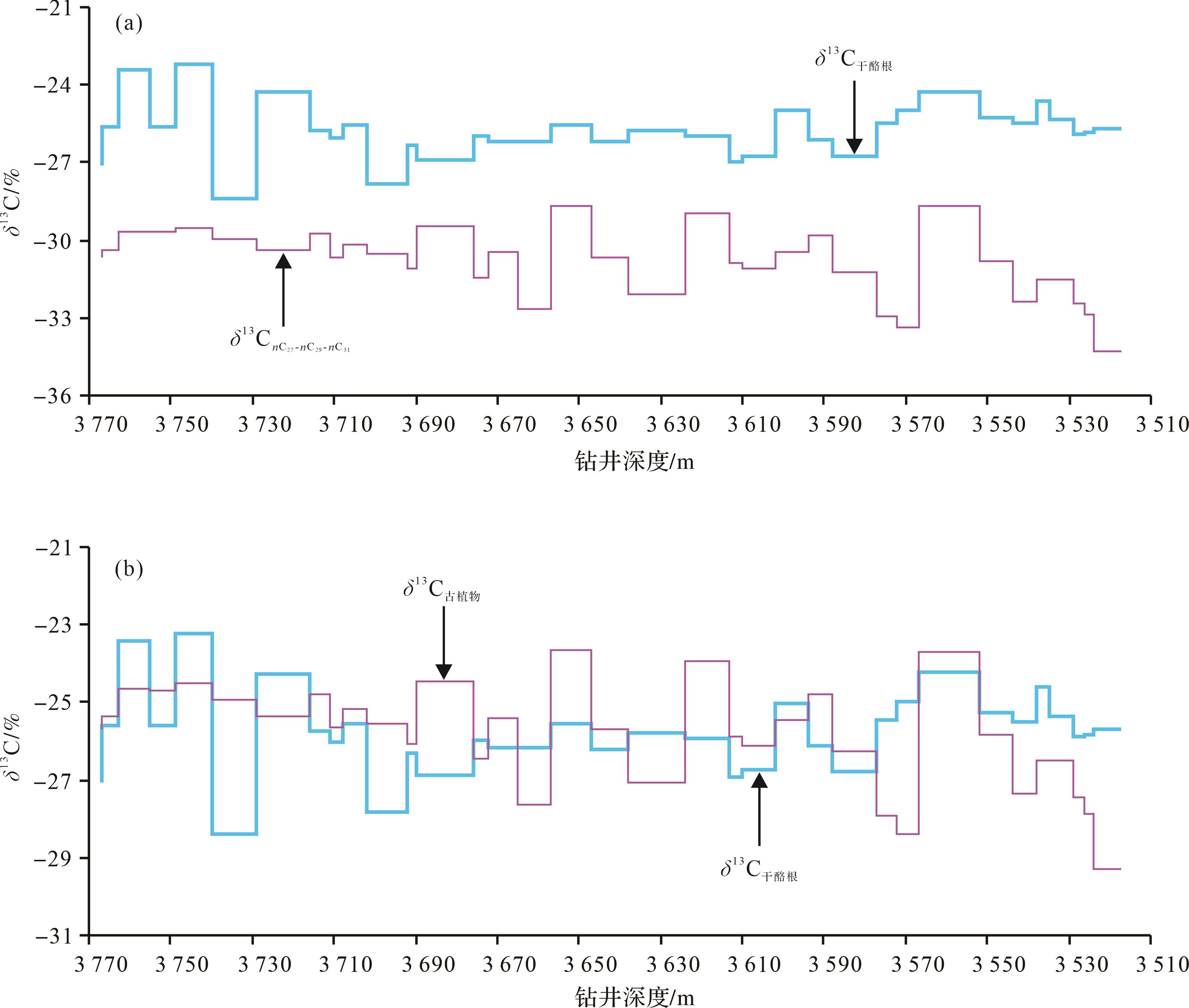
狮68井δ13C干酪根与δ13C nC27-nC29-nC31平均值在钻井剖面的变化显示(图6a),δ13C干酪根与δ13C nC27-nC29-nC31正负向偏移并不同步,这因为岩石中分离的干酪根样品既含有来源于陆生植物器官组织演化而来的镜质组、惰质组、壳质组,还含有一定比例的菌藻类成因腐泥质组分,而正构烷烃是植物叶片表面的蜡质中最常见的组分[23,43-44]。同时,根据本文提出的校对值计算的δ13C古植物变化曲线也与湖相δ13C干酪根有所差异(图6b),δ13C古植物与δ13C干酪根的高低值段变化并不能一一对应,如在井深3 716~3 577 m,δ13C古植物较δ13C干酪根偏正,而在钻井剖面底部3 767~3 716 m和顶部3 577~3 517 m δ13C古植物较δ13C干酪根偏负。这是因为岩石中分离的干酪根既包括湖泊周缘陆地来源的高等植物,也有湖水中生长的水生植物和菌藻类原地沉积的有机质,其δ13C干酪根值至少是三者按不同比例混合的结果。同时,在岩石干酪根显微组分的鉴定中,一般包括腐泥组分和腐殖组分,还可以检测到镜质组、惰质组和壳质组,因此δ13C干酪根的变化也可能与这些显微组分比例大小有关。但是陆地植物特征的长链奇数碳正构烷烃单体分子碳同位素(如δ13C nC29)或δ13C nC27-nC29-nC31平均值则不受有机质组分混合的影响,它直接反映陆地地面环境变化要素(如气温、降水量)的变化。

Figure 6. Carbon isotope variations and comparison of δ13Ckerogen and δ13C nC27-nC29-nC31 (a), δ13Ckerogen and δ13Cancient plants (b)at depth in well Shi 68
目前在研究环境因素如大气CO2浓度、年平均降雨量(MAP)和年平均气温(MAT)影响陆地植物δ13C变化时,所依据的材料是区域和全球现代陆生植物叶片的碳同位素分布数据,因此在引用现代陆地植物δ13C与环境要素定量关系式重建古气候和古环境时,采用δ13C nC27-nC29-nC31平均值可以更精细地描述古环境变化的过程,进而通过对比研究,解释驱动古环境变化的机制。
-
古植物叶片化石的δ13C值可能记录了地质时期生长的植物叶片的碳同位素变化[6⁃7]。但是植物叶片化石在一般陆相沉积中常见而海相地层很少发现,同时这些植物化石叶片至少经历了微生物降解和成岩期热裂解两个阶段的变化过程,其δ13C分析值是否能作为古植物生长期原始叶片的碳同位素信号还需要对比验证。
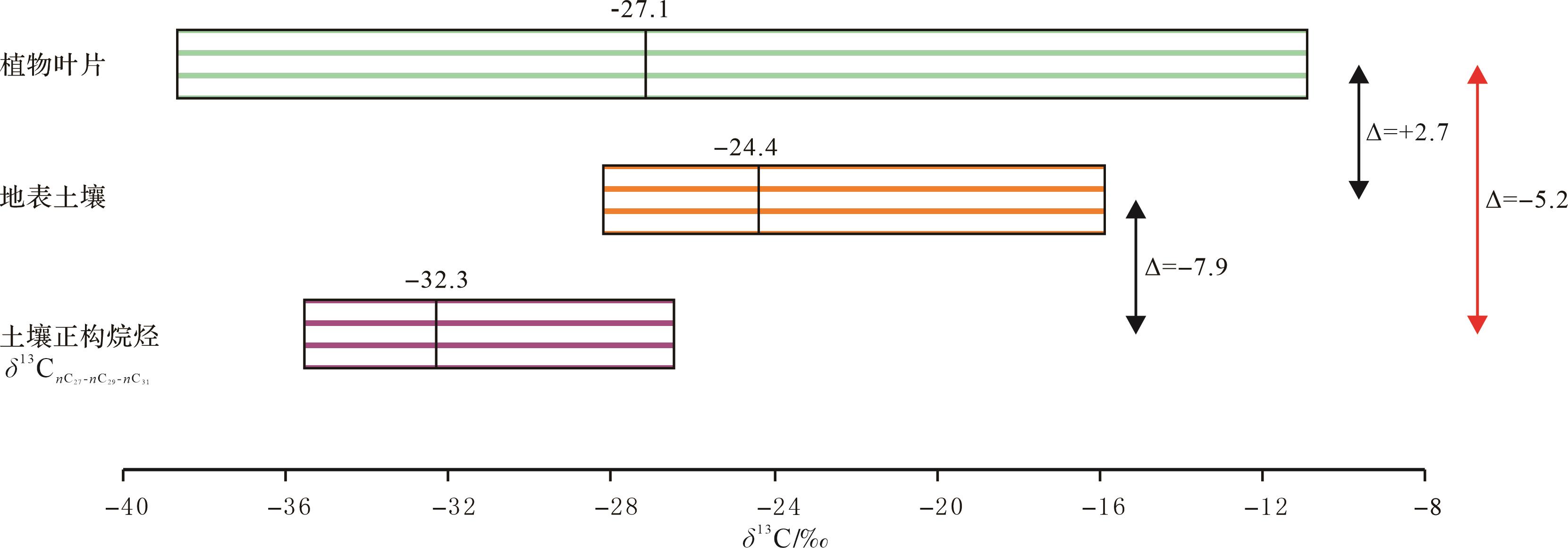
古植物叶片在河流和海浪搬运进入沉积埋藏或成岩阶段之前经历了古陆地面风化降解过程,因此有必要查明沉积埋藏之前其在陆地面古土壤阶段的变化。据全球统计资料,现代全球植物δ13C叶片平均值为-26.8‰,地面土壤有机质碳同位素(δ13C土壤)平均值为-23.1‰,δ13C土壤较δ13C叶片偏正+3.8‰[45]。中国C3型陆生植物δ13C叶片平均值为-27.13‰[45⁃46],Rao et al.[47⁃48]报道的华东和华北现代δ13C土壤平均值为-24.4‰,δ13C土壤和δ13C叶片差值为+2.8‰。根据Rao et al.[47⁃48]数据统计,中国东部现代土壤样品抽提的可溶有机质中δ13C nC27-nC29-nC31平均值为-32.3‰,较中国现代C3植物δ13C叶片偏负-5.2‰(图7)。同时,依据美国Bighorn盆地的资料[49⁃50],该盆地古近纪植物化石层δ13CTOC平均值为-26.0‰,δ13C nC27-nC29-nC31的平均值为-31.1‰,δ13C nC27-nC29-nC31与δ13CTOC的差值为-5.1‰。柴达木盆地侏罗纪大煤沟组δ13C nC27-nC29-nC31-δ13CTOC=-5.3‰和古近纪下干柴沟组δ13C nC27-nC29-nC31-δ13C干酪根=-5.1‰的分馏差值与这两个研究实例的计算结果极为相近(图2,5)。

Figure 7. Carbon isotope distribution and comparison of plant leaf and soil n⁃alkanes in China
-
现代植物叶片中正构烷烃的碳数分布和比值是研究和重建古植被和古气候变化的重要参考指标[51]但是有研究发现植物根、茎、花、果等组织器官也含有一定量的正构烷烃。Fang et al.[52]发现植物花朵要比植物叶片的正构烷烃(nC25-nC33)含量高,Liu et al.[53]证实沉积土壤样品中共计有12%的正构烷烃(nC27-nC33)是植物根系贡献的。Gamarra et al.[54]发现欧洲草本植物不但显示长链奇碳数正构烷烃优势分布,而且植物叶片、叶鞘、花序、茎秆和根系等组织都检测到一定量的正构烷烃。这就说明沉积地层中检测到的指示陆生植物来源的长链奇数碳正构烷烃nC27、nC29和nC31为植物组织混合来源,而且这些组织器官的碳同位素分布还存在一定差异[55⁃56]。因此,沉积地层中检测到的正构烷烃δ13C nC27-nC29-nC31是古植物组织的混合信号,而不是单一古植物叶片碳同位素值。
陆地植物有机组织和陆表土壤有机质在埋藏期,随着温度升高和压力增大,碳同位素成分可能发生变化。基于沉积有机质的时代分布和实验室热模拟实验研究,一般认为植物化石和岩石中有机质随着成岩强度增加,δ13C明显偏向重碳同位素富集的方向演化[57⁃58]。然而,Tian et al.[59]热解实验结果表明,在实验模拟温度升高至300 ℃时,即对应镜质体反射率Easy Ro变化自0.48%升高到0.71%时,δ13C nC27-nC29-nC31的碳同位素仅偏正0.80‰,这意味着采用δ13C古植物=δ13C nC27-nC29-nC31+5.0‰可适用于未成熟到低成熟阶段古植物叶片的复原计算。
4.1. 沉积有机质干酪根与陆生植物正构烷烃的碳同位素分布特征
4.2. 钻井剖面沉积有机质干酪根与陆地古植物的碳同位素变化特征
4.3. 陆生植物与陆表土壤以及沉积有机质碳同位素变化的对比分析
4.4. 陆地植物组织器官碳同位差异及埋藏成岩过程有机质碳同位素变化的对比分析
-
(1) 植物化石不同组织器官样品δ13C测量值的误差。在现代植物碳同位素值研究工作中,一般通过测量植物叶片的碳同位素值研究木本植物、草本植物和陆地C3植物、C4植物的变化范围,进而研究纬度和高程变化以及地面温度高低、降雨量大小对植物δ13C的影响。但在古植物碳同位素研究中,因为叶片化石稀少,有时通过采集植物茎秆、树皮或植物组织残存碎片测量的δ13C表征古植物的同位素值[8,60],煤层和木化石的δ13C也是常见的指示陆地植物碳同位素的指标[61⁃62]。前人研究发现,植物根系、树干和纤维素的δ13C较植物叶片偏正[43,63⁃64],这可能造成δ13C古植物计算值的误差。
(2) 干酪根显微组分含量相对变化对δ13C分析值的干扰。陆相沉积地层的煤层、古土壤有机质和岩石分离的干酪根δ13C值,可能受菌藻类成因的腐泥质组分相对含量大小的影响。一般煤岩和干酪根显微组分可划分为镜质组、惰质组、壳质组和腐泥组四部分,据Whiticar[65]研究成果,惰质组δ13C偏正,δ13C平均值为-23.4‰,壳质组δ13C偏负,δ13C平均值为-23.9‰,但腐泥组偏负可达-24.3‰,误差约为±1.0%。另外,柳益群等[66]和解光新等[67]研究发现,腐殖煤δ13C值为-24.4‰,腐泥煤δ13C值偏负可达-32.2‰,二者相差接近±8‰。由于陆相沉积干酪根组分中或多或少都含有一定量菌藻类成因的腐泥质组分,这对采用δ13C干酪根作为陆地高等植物δ13C的替代指标提出了挑战。
(3) 古代岩石样品与现代陆生植物中正构烷烃的碳数分布和δ13C值的差异。现代植物叶片正构烷烃的碳数分布范围为nC21~nC35,奇碳数和偶碳数丰度差异明显,并以出现高含量长链正构烷烃nC27、nC29和nC31为特征峰[23, 51],木本植物主峰碳为nC27或nC29,草本植物主峰碳为nC31。在地质样品中,长链正构烷烃的奇偶优势减小乃至消失,奇碳数和偶碳数正构烷烃含量几乎相等。但从正构烷烃单体分子碳同位素分布来看,奇偶碳数正构烷烃nC26~nC32与奇碳数正构烷烃nC27~nC31碳同位素平均值十分相近,分别为-29.5‰和-29.8‰。
(4)植物化石与现代裸子植物和被子植物正构烷烃 δ13C nC27⁃nC29⁃nC31 对比分析。Diefendorf et al.[68]研究表明,现代C3型植物δ13C叶片与正构烷烃nC27、nC29、nC31碳同位素(δ13C正构烷烃)平均差值分别为-4.6‰、-5.6‰、-6.0‰,这其中C3型裸子植物中乔木植物δ13C叶片与正构烷烃nC27、nC29、nC31碳同位素平均差值分别为-4.1‰、-4.2‰、-4.5‰。C3型被子植物中乔木植物δ13C叶片与正构烷烃nC27、nC29、nC31碳同位素平均差值分别为-4.3‰、-5.2‰、-5.6‰,C3型被子植物中草本植物δ13C叶片与正构烷烃nC27、nC29、nC31碳同位素平均差值分别为-4.8‰、-6.0‰、-6.3‰。柴达木盆地古近纪含化石岩石δ13C nC27-nC29-nC31平均值较δ13C古植物偏负-4.7‰,但样品数量少,它是否具有普遍意义需要进一步研究。侏罗纪δ13C古植物与δ13C nC27-nC29-nC31的平均差值为-5.1‰,这一差值小于现代C3型裸子植物的δ13C nC27-nC29-nC31平均值,但大于现代C3型被子植物的δ13C nC27-nC29-nC31平均值。
(5) 复原古植物碳同位素δ13C的校对公式适用条件。钟建华等[69]研究发现,柴达木盆地新生代平均地温梯度2.8 ℃/100 m,干酪根镜质体反射率(Ro)与钻井井深(D)之间具有如下关系:Ro(%)=0.000 1D+0.443 2。根据狮68井采样深度计算,狮68井下干柴沟组镜质体反射率Ro(%)为0.8%,对应中成岩阶段。舒豫川等[70]实测英西地区下干柴沟组样品Ro介于0.45%~0.63%,平均为0.56%,有机质演化进入成熟期。柴北缘侏罗纪中统大煤沟组煤岩组成以亮煤为主,暗煤次之,煤岩样品的Ro平均值为0.6%[71]。侏罗系泥页岩Ro变化范围为0.52%~1.43%[72],有机质处于低成熟到高成熟阶段。本文研究的样品涵盖了成岩作用跨度从浅埋藏到深埋藏区间,因此我们提出的通过高碳数长链正构烷烃的校对和复原陆生古植物碳同位素的参数,即δ13C古植物=δ13C nC27-nC29-nC31+5.0‰,适用于沉积地层有机质热演化程度处于未成熟阶段到低成熟阶段的样品计算校对。
(6) 计算和复原古植物δ13C古植物参数的对比研究。Bourque et al.[73]在研究白垩纪和古近纪δ13C古植物分布时,采用Chikaraishi et al.[74]实验结果,按下式校对计算古植物的碳同位素值:水生植物δ13C植物叶片(‰)=δ13C nC25+7.4‰,陆生C3型被子植物δ13C植物叶片(‰)=δ13C nC29+3.1‰,陆生C3型裸子植物δ13C植物叶片(‰)=δ13C nC29+4.7‰。Schlanser et al.[75]在重建白垩纪—古近纪古植物δ13C植物叶片变化时,引用Diefendorf et al.[68]的研究资料,按下式校对计算木本植物的碳同位素值:δ13C木本植物(‰)=δ13C nC29+5.2‰,δ13C木本植物(‰)=δ13C nC31+5.6‰,而草本植物碳同位素值的校对值取δ13C草本植物(‰)=δ13C nC29+6.0‰,δ13C草本植物(‰)=δ13C nC31+6.3‰。应该说明,这两个研究实例关于古植物与正构烷烃碳同位素的分馏差值是基于现代植物分析资料,并且假定古植物叶片和正构烷烃在埋藏成岩过程δ13C无明显富集和亏损。本文根据经历了表生成土期有机质细菌降解作用和埋葬成岩期热解生排烃等复杂地质过程的植物化石和正构烷烃碳同位素实测值提出了δ13C古植物校对计算值,可能更适用于地质时期δ13C古植物的复原和重建。
-
(1) 柴达木盆地侏罗纪中统大煤沟组煤系地层和古近纪下干柴沟组的植物叶片化石和少量植物茎秆化石δ13C古植物介于-26.2‰~-22.7‰,平均值为-23.7‰,记录陆地高等植物叶片的δ13C nC27-nC29-nC31介于-30.2‰~-27.5‰,平均值为-28.7‰,植物叶片化石δ13C古植物与正构烷烃δ13C nC27-nC29-nC31差值-5.0‰。
(2) 狮子沟地区狮68井下干柴沟组与盐湖相蒸发岩互层产出的富有机质黑色泥岩样品δ13C干酪根介于-28.4‰~-23.2,平均值为-25.8‰,表征陆地高等植物来源的δ13C nC27-nC29-nC31介于-34.3‰~-28.7‰,平均值为-30.9‰,δ13C干酪根与δ13C nC27-nC29-nC31差值为-5.1‰。
(3) 在沉积地层有机质碳同位素研究中,考虑到古植物叶片和茎秆δ13C的差异以及埋藏成岩的影响,本文提出可采用5.0‰作为δ13C nC27-nC29-nC31平均值对陆地古植物δ13C古植物的校对值,即δ13C古植物=δ13C nC27-nC29-nC31+5.0‰,它可以更准确地标定古大气CO2浓度、年平均降雨量等古气候要素的变化过程,减少全岩有机碳和干酪根碳同位素受到多组分混合作用的影响和干扰。

 DownLoad:
DownLoad: