

 下载:
下载:


-
微生物岩是一种与微生物席密切相关的沉积记录,包括由微生物席内新陈代谢活动产生的有机—无机矿物,以及由微生物席捕获、粘结席外界颗粒形成的特殊沉积构造,可根据相关宏观特征进一步分为叠层石、凝块石、树形石、均一石、核形石和纹理石[1⁃6]。其中叠层石是分布最广且研究最为详细的类型,而凝块石(Thrombolite)一词则缘自20世纪60年代。1967年,Aitken首先将加拿大南部落基山地区寒武统—下奥陶统地层中具有聚集状凝块(Clot)结构的岩石命名为凝块石(Thrombolite),并指出其与叠层石相比缺乏连续状纹层构造[7⁃8]。Riding则进一步认为凝块石主要由颜色、结构迥异的凝块和基质构成,亦可包含除凝块之外其它组分[1⁃4]。整体来说,凝块石具有广阔的时空展布范围。就分布时代而言,凝块石多分布在晚新元古代至早奧陶世的生物建隆中,之后受后生动物分化影响,其分布丰度锐减并多局限在与极端环境相关的地层中,如志留纪早期、泥盆纪末期、侏罗纪早期等生物大灭绝之后,以及第三纪盐度事件期间;就产出环境而言,凝块石可分布于局限港湾、开阔潮下带、淡水湖、超盐湖、碱水湖以及热泉等环境[1⁃10]。
凝块石成分复杂,分布宽泛,缘自其本身的成因多样性,同时也从侧面说明凝块石的研究存在着不小的难度。鉴于凝块石的整体展布不及叠层石广泛,加之经常和其他类型的微生物岩呈共生—过渡关系,难以区别开来,因此前人多将凝块石放入微生物岩组合中进行描述。如Feldmann和Planavsky认为现代巴哈马的叠层石—凝块石组合缘自被后生动物破坏的叠层石,同时还包括准同生矿化、次生胶结和局部碳酸盐溶解作用的影响[9⁃10],Dupraz et al.[11]则认为真核生物的繁殖才是破坏叠层石的主要元凶;Tang et al.[12-13]通过分析华北地台前寒武纪叠层石—凝块石组合,认为叠层石与凝块石分别由丝状细菌和球形细菌构建而成;Riding et al.[1⁃4]则认为丝状菌可瓦解叠层结构并促进凝块组构的发育。综上我们可以看到,相关组合研究多集中在“凝块石—叠层石组合”方面,而对于成分更复杂、非均质性更强的凝块石,却鲜有研究思路和相关实例。
寒武纪生命大爆发(Cambrian explosion)后,微生物岩受微生物和后生生物的综合影响,其结构与组成明显复杂于前寒武纪微生物岩。近几年来,有学者提出引用“微生物席生态系统矿化作用”的概念和理论进行进一步的研究。目前与微生物席相关的研究实例和培育实验,主要为复杂微生物岩的形成提供了四点新思路,包括:1)微生物席内存在球状原生白云石,可视为沟通普通沉积作用和微生物活动的桥梁[14⁃21];2)与薄微生物席相比,厚微生物席自下而上存在着明显的分异性,主要表现为微生物群落以及相关新陈代谢方式的变化[14⁃21];3)薄微生物席可形成叠层结构清晰的叠层石,而厚微生物席中,生物成因的多孔状微组构可粘附方解石/白云石沉淀物,进而形成凝块并构成粗糙纹层状至不规则状的微生物岩[14⁃21];4)球状—丝状蓝细菌或其他可进行有氧光合作用的生物,其单独或协调活动可促进席内矿化作用和碳酸盐沉淀[22⁃25],如活跃的有氧光合作用消耗CO2,导致pH值升高,导致结晶较差的Mg⁃Si泡状细胞胞外有机质沉淀,以及蓝细菌和胞外有机质降解,可经硫酸还原菌(SRB)诱导成核[26⁃35]。在以上理论指导下,梅冥相等[15]曾对位于辽东半岛本溪田师傅剖面和大连金州湾剖面的长山组凝块石进行剖析,为微生物席内复杂的钙化和沉积现象提供了典型实例。然而若将微生物席内部视为一种独特的微环境,将微生物席内沉积看做一个年轻的沉积学分支[16⁃17],那么在研究微生物席与复杂的微生物岩时,其与大沉积环境的互动也应被考虑进去。
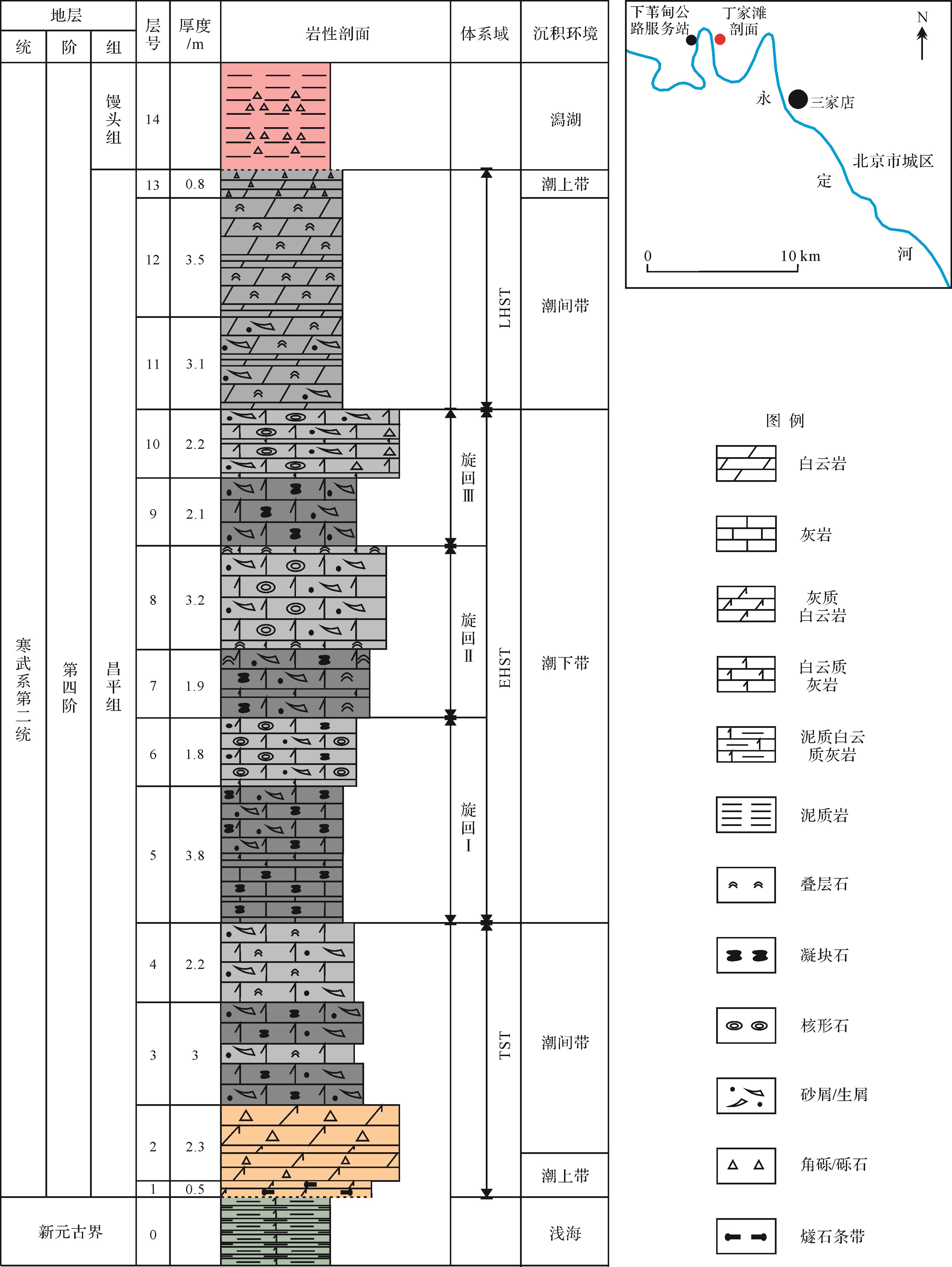
在北京市门头沟区,下苇甸地区的众多寒武系剖面作为典型教学剖面,其相关岩相组合、沉积环境以及地层划分对比特征得到了较为充分的研究。在该地区第二统昌平组底部,发育于含燧石角砾白云岩之上的一套豹斑灰岩可与周口店地区黄院东山梁239高地的下寒武统府君山组豹斑灰岩相对应[18⁃19]。然而经过实地踏勘,笔者发现该套豹斑灰岩含有大量微生物成分,实则为一套沉积在高能环境下的凝块石,若单纯套用经典豹斑灰岩的相关定义,恐怕不妥。因此,笔者选取下苇甸地区丁家滩剖面,重点探讨了第二统昌平组凝块石的特点和成因,最后以该段岩层为例,对凝块石和豹斑灰岩的区别进行探讨,力求为“寒武纪生命大爆发(Cambrian explosion)”之后的凝块石研究提供一个典型的分析实例,有助于恢复“奥陶世生物大辐射事件(Ordovician radiation)”之前的古地理环境,重塑“后生动物先驱阶段(Pioneer metazoan stage)”的地球演化史[22]。
-
华北地台在青白口系景儿峪组沉积之后,经过近4亿年的剥蚀作用形成了一个“巨型不整合面”,与此同时华北地台北缘拉张形成古亚洲洋,之后的海侵事件导致不整合面上沉积了分布广泛的昌平组碳酸盐岩[23⁃25]。昌平组富含三叶虫、腕足类、软舌螺等化石,自下而上可包括3个三叶虫带:Megapala⁃ eolenus,fengyangensis, Redlichia sp.。由于三叶虫和牙形石等年代指向性化石具明显穿时性,因此华北地台的寒武系地层系统多建立在岩石地层学的基础上,即组大致与阶对等,且地层普遍存在着跨统现象。在寒武系四分框架中,昌平组大致位于第二统第四阶下部[36⁃40],底界年龄约515 Ma,而顶界年龄分歧较大,多集中在509 Ma左右[21⁃30]。
丁家滩剖面(40°0′1.07″ N,116°02′13.35″ E)位于北京市门头沟区109国道边,西距下苇甸公路服务站约2 km,主要出露元古宙—古生代地层。其中昌平组厚约30 m,整体对应一个三级层序,包括海侵体系域(TST)、早期高位体系域(EHST)和晚期高位体系域(LHST)[19](图1),其底部以I型层序界面与下伏景儿峪组相隔,发育一套底砾岩;下部TST厚约8 m,主要由潮间带凝块石和叠层石组成,见流水波痕;中部EHST厚约15 m,整体砂屑含量较高,见大量生物潜穴和生物扰动构造,主要由沉积在潮下高能环境的凝块石和核形石组成,可构成3个旋回;上部LHST厚约7 m,主要由叠层石构成,大致对应潮间—潮上带,指示沉积环境整体变浅。昌平组顶部厚达数厘米至数十厘米的古喀斯特角砾透镜体/层指示长时间暴露,代表I型层序界面[19,24]。
-
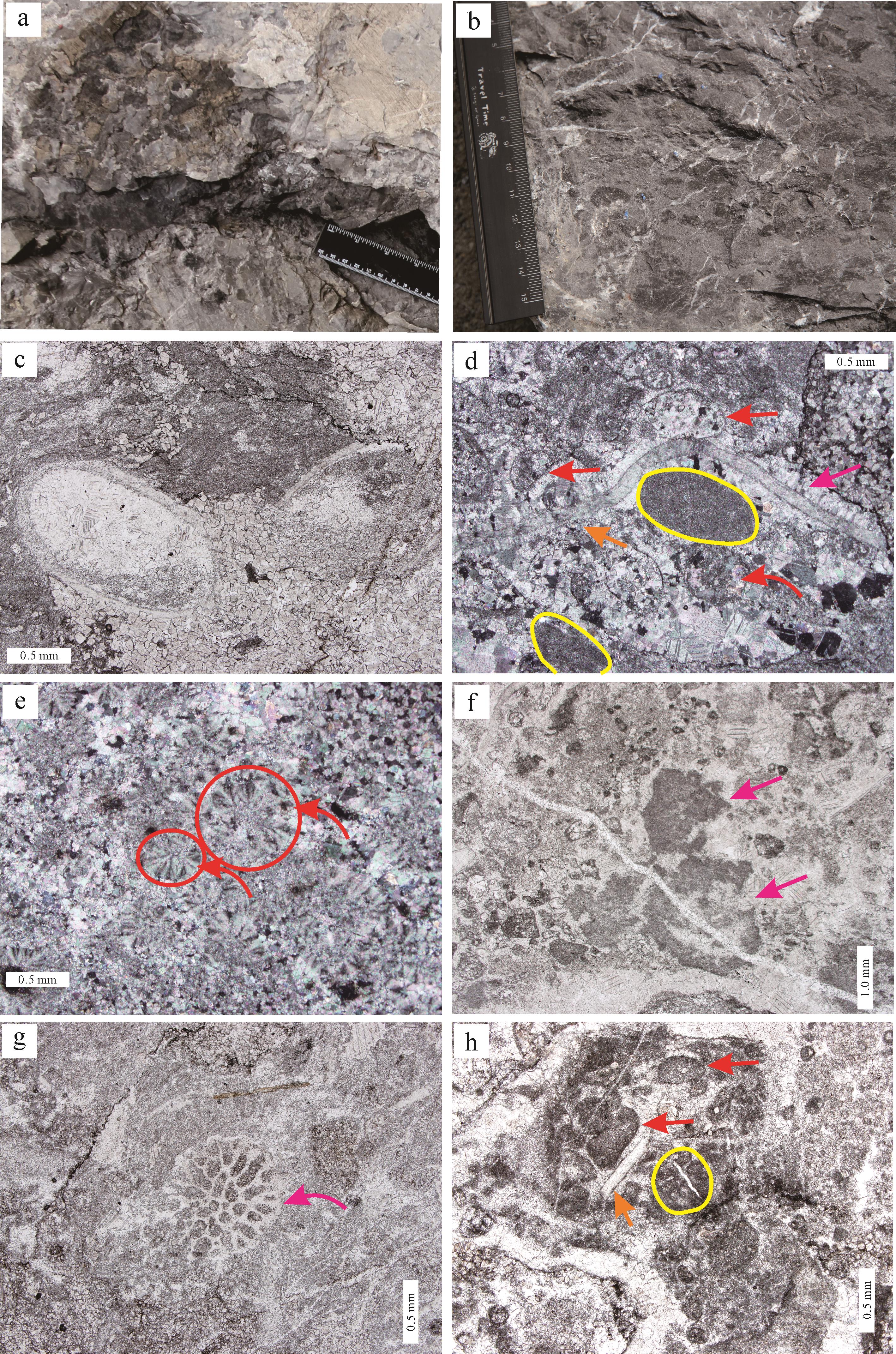
昌平组TST和EHST中可见多套层状凝块石(图2a),可与层状叠层石共生(图2b,c),其单层厚度多为米级,整体起伏较小,横向延伸可达数米至数十米,一般与周边宿主岩石(多为产自高能环境的砂屑灰岩,或泥质含量很高的泥质灰岩,部分可具叠层构造)具有明显的岩性差异,巨观上符合“生物层(Biostrome)”的定义[4]。宏观上可见明显的暗色凝块和浅色基质。笔者从凝块的形态出发,将凝块石分为斑状凝块石、带状凝块石和网状凝块石3类。其中斑状凝块石可归为Riding分类中的粗粒黏结凝块石(Coarse agglutinated thrombolites),而带状凝块石和网状凝块石属于次生凝块石(Post⁃depositional thrombolites)[1⁃4]。

图 2 北京西郊寒武系第二统昌平组生物层与凝块石
Figure 2. Biostromes and thrombolites in the Cambrian Changping Formation, Series 2, western Beijing
-
斑状凝块石主要见于EHST,为昌平组最常见的凝块石。宏观上其风化面主要呈黄色、红褐色,斑纹结构不甚清晰(图3a),新鲜面可见其由浅色基质和不规则暗色凝块组成(图3b)。其中基质多以灰质为主,主要由微晶方解石构成;而白云质凝块多呈杂乱状分布,大小介于0.5 cm×0.8 cm至3 cm×7 cm,含量约占整体岩石的30%~50%,镜下见大量细晶—粉晶白云石(图3c),可包含两种显微凝块结构。

图 3 斑状凝块石特征
Figure 3. Characteristics of porphyritic thrombolites
第一种显微凝块包含大量颗粒物质,如砂屑、三叶虫碎片和鲕粒,可见少量泥晶物质(图3d)。这些弥散状暗色泥晶可能为微生物及胞外聚合物(EPS)降解的残余物质,或诱导碳酸盐矿物沉淀的产物(多由微生物钙化机制引起的局部pH值和
鲕粒(直径多<0.2 cm)主要包括三种:1)馒头状鲕粒(图3d),多为偏心结构。核心多位于平直端一侧,主要由弥散状云雾泥晶构成,可能代表微生物活动的副产物[41⁃42];凸起端一侧圈层局部可见黑色小颗粒,可能为微生物钻孔活动的残余物质[43⁃45],因此与砂屑相比,该种鲕粒亦可被理解为受微生物改造的微生物成因微晶灰岩碎片。鲕粒凸起端上部可见马牙状胶结物,而平直端则几乎不发育,加之平直端多依靠在三叶虫碎片或者其他颗粒上,指示馒头状鲕粒可能具有附着性质[46]。2)同心圆状鲕粒,多呈零星状分布,一般具有核心,多和馒头状鲕粒共生。整体圈层数目较少,部分甚至与皮壳结构、皮壳粒(Cortoid)类似。外部一般发育较厚的富有机质暗色泥晶套,可与芙蓉统凤山组发育的底栖有核放射鲕相互对照[47]。3)放射状鲕粒(图3e),多呈聚集状,大多具有放射状结构,可能指示其初始成分为低能环境下的文石。参照Gerdes的现代微生物席培养实验[46],以及梅冥相基于北京西郊寒武系芙蓉统凤山组生物层的研究成果[37],可认为这三种低能鲕粒均形成于厚微生物席内部,因此有悖于整体高能大环境。它们的存在也代表了席内的复杂钙化作用,更反应了席内的非均质性和多样性,从而导致凝块石具有复杂的内部组成[41,47⁃49]。
第二种显微凝块主要由大量泥晶物质和部分颗粒共同组成。镜下团状泥晶物质可具不明显包壳(图3f),亦可见与团状泥晶物质整体外形相似的单个绿藻节片,细节上呈藕片状—管孔状(图3g),这可能说明团状泥晶物质来源于绿藻节片的直接降解,但不排除微生物及EPS的降解作用及诱导沉淀作用。若从整体来看,可见厚微生物席的互动形式和组成成分有变复杂的趋势[16⁃17]。颗粒多集中分布,主要包括砂屑和生物碎片,一般由大量泥晶物质粘结而成(图3h)。富含破碎状泥晶物质的砂屑与破碎状生物碎片共生进一步指示了微生物作用和动荡水体共存的现象。
-
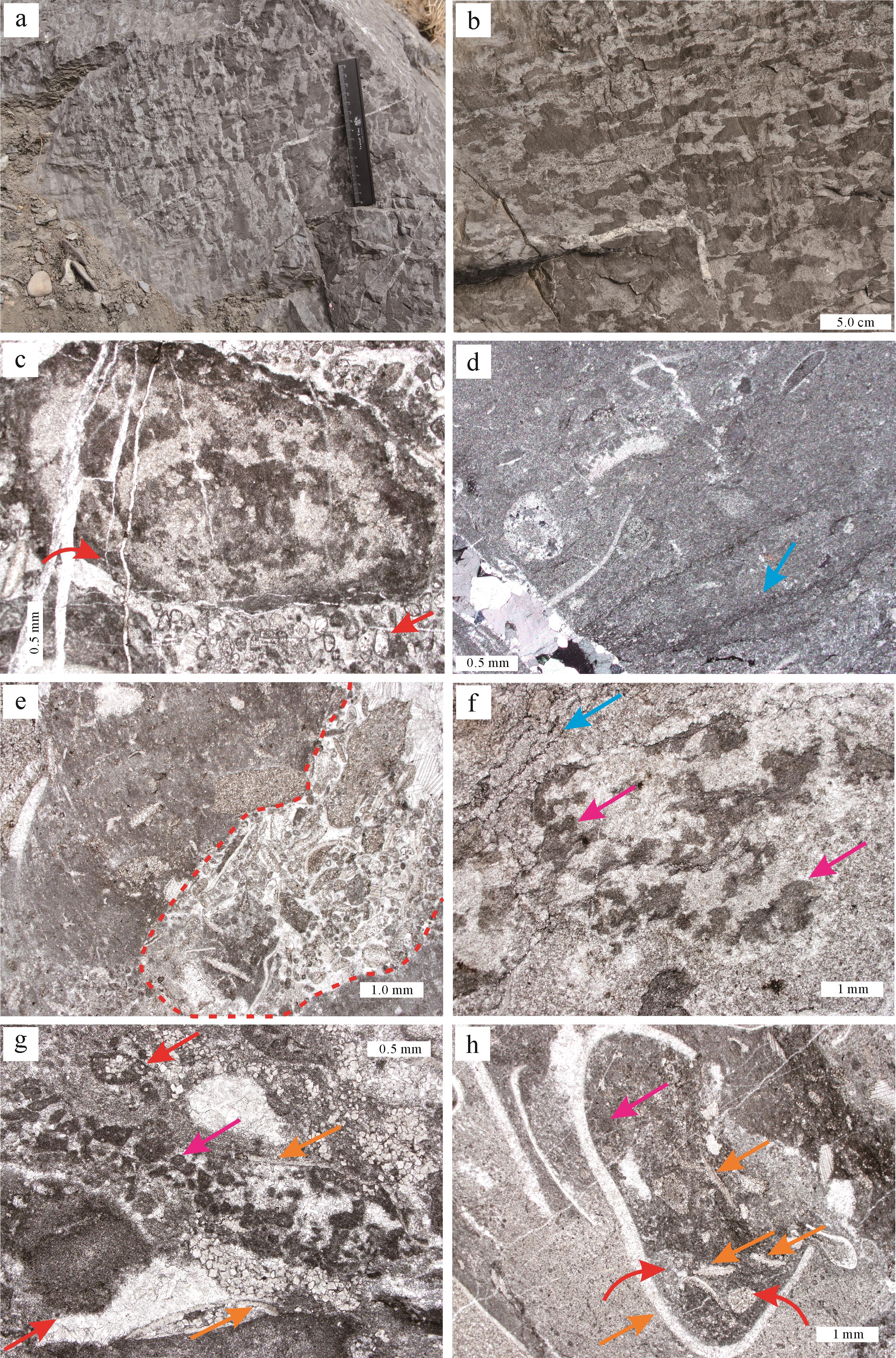
带状凝块石主要见于昌平组TST下部,多与叠层石存在着侧向或垂向渐变关系(图4a)。宏观上风化面一般呈浅灰色,新鲜面呈灰色,见大量钻孔结构,以及深色带状凝块和浅色基质。其中云质带状凝块大多介于0.6 cm×0.1 cm至2 cm×7 cm,含量约占整体岩石的40%~60%,而基质和叠层结构多由灰质组成。部分带状凝块自边缘向上翘起,与泥裂构造较为相似,指示潮间带间歇性暴露特点[50];有时亦可呈撕裂状(图4b),凝块之间及附近多发育凹凸不平的冲刷面和水流波痕,以及少量毫米级核形石和砂屑(图4c),代表了潮间带往复性强水动力特征[51]。

图 4 带状凝块石特征
Figure 4. Characteristics of striped thrombolites
显微镜下可见基质中发育弥散状泥晶显微叠层结构(图4d),垂直纹层分布的钻孔多被大量生屑、砂屑及显微凝块结构充填(图4e),也可逐渐过渡为含大量生屑、砂屑的粗粒显微叠层结构。后者中可见两种显微凝块结构:第一种主要由弥散状泥晶构成,可逐渐过渡为微生物纹层(图4f);第二种主要由团块状泥晶和包壳颗粒组成,多具粘连呈席的趋势(图4g)。组成带状凝块的显微凝块结构与钻孔中的显微凝块结构类似,主要以生物碎片为格架,内部被大量泥晶、馒头状鲕粒以及生屑充填(图4h),为典型厚微生物席内产物。
-
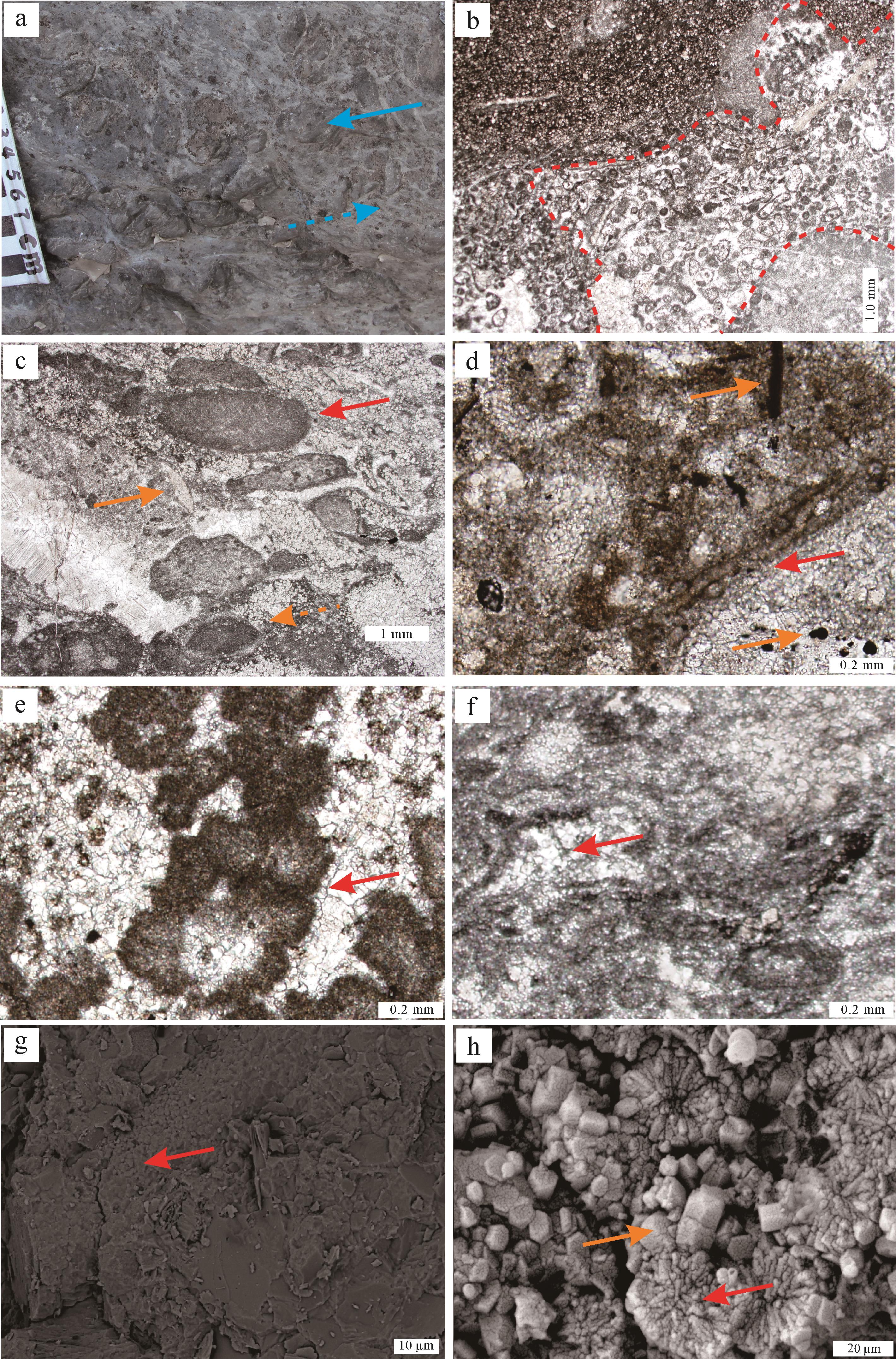
网状凝块石主要见于昌平组TST上部。宏观上其风化面呈中灰色,新鲜面主要呈深灰色,可见深色凝块主要呈不规则网格状,网格大小0.7 cm×1.0 cm至3 cm×4 cm,主要表现为暗色凝块被封闭在浅色基质之间,其中凝块含量约占整体岩石的60%~80%,多为云质,而基质则可呈灰质—云质(图5a)。镜下见深色凝块主要由显微凝块结构组成,多与不规则洞穴伴生,疑似大生物挖掘、扰动痕迹(图5b)。其中洞穴内部见大量生屑、砂屑、泥晶物质以及显微凝块结构,与带状凝块石较为类似,具有明显的厚席特征。

图 5 网状凝块石特征
Figure 5. Characteristics of reticular thrombolites
基质中可见少量显微凝块结构,主要由核形石雏形、砂屑、生屑、泥晶物质组成(图5c)。泥晶物质中可见管状葛万菌(Girvanella)和阿哲菌(Izhella)。这些具有光合作用的蓝细菌整体保存完好,结构清晰,为蓝细菌鞘活体钙化作用的产物[52]。其中单体葛万菌呈管状,直径大多集中在20 μm左右,管壁多由暗色泥晶物质组成,内部则由亮晶方解石/白云石充填,见黄铁矿化菌丝与粒状黄铁矿与之伴生,可能指示硫酸还原菌(SRB)的活动(图5d);阿哲菌集合体多呈西兰花状共生,单体多由泥晶物质构成,中心可被亮晶方解石/白云石充填,横切面可呈空心花朵状(图5e),和肾形菌(Renalcis)相比其菌壁更厚,且菌壁并不分层[53⁃54]。另外,核形石雏形的圈层中可见窗格状孔隙,多分布在丝状菌(葛万菌?)缠绕而成的网状体中(图5f),可能为光合作用产生的氧气气泡残留[49]。扫描电镜显示核形石内部发育大量直径为20~30 μm的蓝细菌菌丝(图5e)。其中内部呈放射状的微米级球状白云石与粒状白云石共生(图5f),可能与中度嗜盐好氧细菌作用有关[55]。
-
露头实测和镜下观察结果表明北京西郊寒武系第二统昌平组凝块石灰岩主要发育在高能环境,这种沉积大环境导致凝块石中含有大量砂屑和生屑成分;其次,TST凝块石多发育在潮间带,而EHST凝块石多发育在潮下带,这种水体逐渐加深的变化导致指示较浅沉积环境的虫迹与叠层石仅仅发育在TST潮间带凝块石中。
另外,昌平组微生物岩中可发育两种微生物席,对应两种截然不同的沉积微环境:第一种为薄微生物席,多形成成层性较好的叠层石,其微生物群落较为单一,仅发育藻类,厌氧微生物和蓝细菌等等[14,48⁃49]。薄微生物席具有捕获粘结作用,可形成粒度较粗的叠层石,且被捕入席中的颗粒增加了微生物席的复杂性,有利于向厚微生物席转化;第二种为厚微生物席,多形成微组分复杂的凝块石。结合近年来针对形成凝块石的微生物席培养试验和现代实例,以及昌平组具体特点,可见这种厚微生物席具有如下特征:1)多由交织成网状的微生物群落(如葛万菌和阿哲菌)组成,席中微生物可形成多孔状—不规则状微组构(如弥散状泥晶物质),并进一步形成粗糙纹层状到高孔隙状的不规则微生物岩(如凝块石)[55⁃57];2)其内部存在多种生物群落,自厌氧、化能自养生物(如SRB)至喜氧、光能自氧生物(如蓝细菌)均有涉及[10,14,55⁃57];3)具有黏性并可捕捉各种类型颗粒(如三叶虫碎片、砂屑等),亦可在内部形成新的矿物和颗粒(如形态各异的鲕粒及各种胶结物等)[55⁃57]。与薄层相比,很明显厚席的沉积微环境对凝块石的成分和基本特征影响更大一些,如斑状凝块石内部可见大量低能底栖鲕粒,这与高能大沉积环境明显不符。
-
以TST带状凝块石为例,其多发育在潮间带,镜下可见带状凝块具典型厚席特征。加之1)翘起的带状凝块边缘指示间歇性暴露环境;2)而撕裂状的带状凝块边缘和指示强水动力条件的核型石、砂屑交互共生;3)凝块间可见明显冲刷痕迹。那么这些长条状凝块则可看做是被失水干裂以及暂时性流水分割成多块的厚席,即沉积大环境可对尚未固结成岩的微生物沉积物进行宏观尺度的改造。值得一提的是,若将水道内部发育的弥散状包壳核形石仍看做是微生物席内的产物,那么厚席也许并不是像普通沉积物那样被水道完全切割开来,而是更具有一种“藕断丝连”的特征。
另外,现代巴哈马凝块石研究表明,凝块石产生于微生物和后生动物共存的混合底栖生态系统。微生物和后生动物一方面通过捕获和被捕获的形式,直接参与微生物席的构成,另一方面通过钻孔作用和挖掘改造作用,对原本形成的微生物岩进行改造[58⁃60]。在带状凝块石和网状凝块石中可见大量垂直纹层的钻孔结构和不规则的洞穴结构,这些钻孔和洞穴内部中仍有显微凝块微组构发育,说明这两种改造作用和微生物席的自身生长可同时进行。因此,微环境中各种生物与原始沉积物的互动,影响了昌平组凝块石的微观特征。
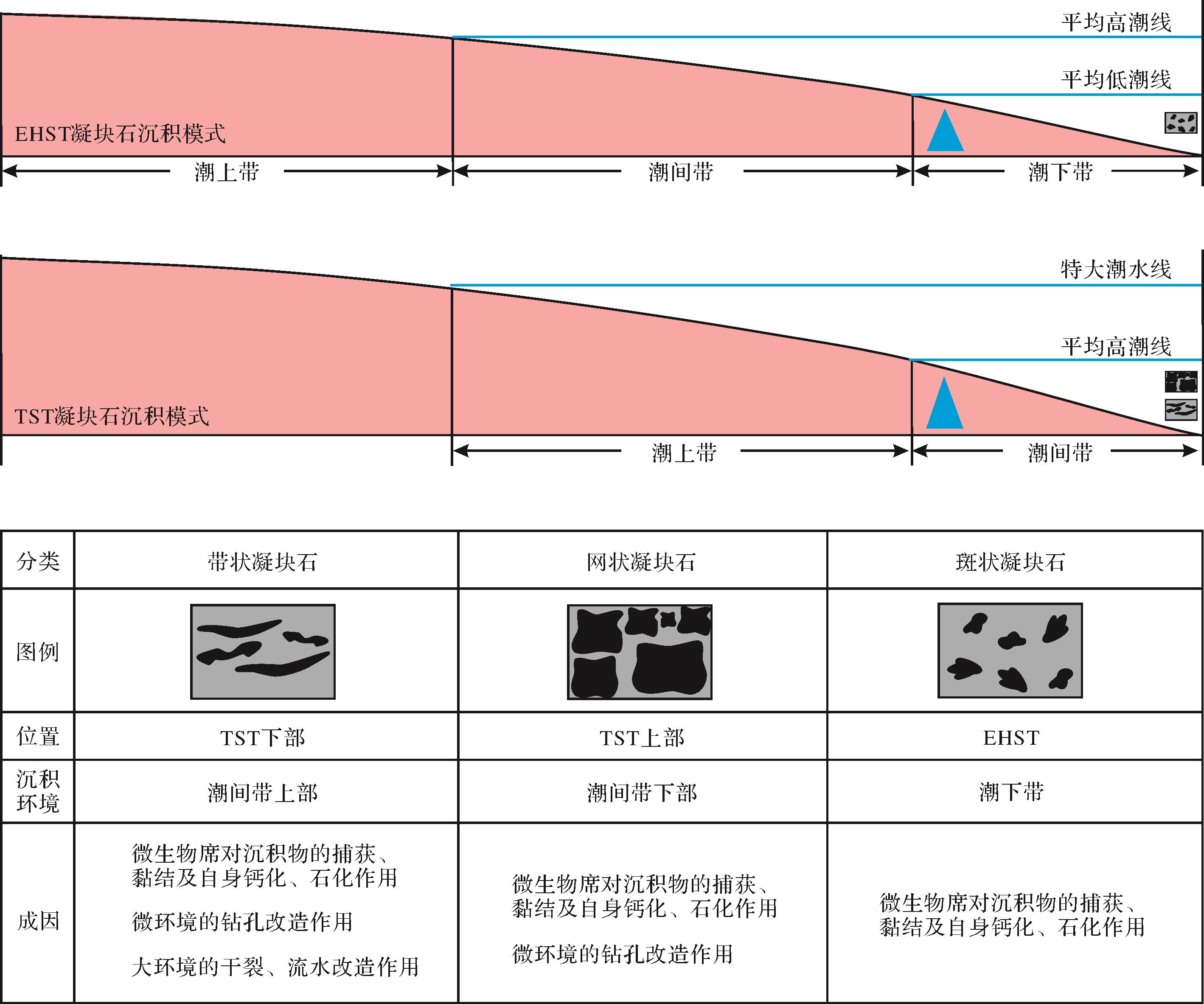
根据以上分析,笔者建立了北京西郊寒武系第二统昌平组凝块石沉积模式(图6):TST时期整体沉积环境为潮间带,水体较浅,氧气光照充足,有利于后生动物和微生物的快速生长,因此除却微生物席对沉积物的捕获、粘结及自身钙化、石化作用外,席内微环境的钻孔改造作用也十分常见。在TST下部,水体更浅的潮间带上部,甚至可以看到失水干裂及暂时性流水将厚席分割成多块。至EHST时期整体沉积环境为潮下带,整体水体能量较高,因此和潮间带相比其席内包含更多颗粒成分,这也能从侧面说明该环境下厚微生物席整体粘性较大,加之过于浑浊动荡的水体不利于生物/微生物的挖掘作业,遗迹不好保存,因此该环境下形成的斑状凝块石基本由微生物席内沉积直接构成。

图 6 北京西郊寒武系第二统昌平组凝块石沉积模式图
Figure 6. Sedimentary model of the Cambrian Changping Formation, Series 2, western Beijing
-
北京西郊寒武系第二统昌平组发育的凝块石具有复杂的结构和成分,反映了“寒武纪微生物岩复苏”以及“新元古代后叠层石衰退”这两个独特的地质现象[1⁃4]。依据本文及前人研究成果,笔者对豹斑灰岩和凝块石的区别进行了进一步的探讨。
豹斑灰岩和凝块石均由不规则豹斑/凝块和基质组成,因此在野外露头鉴定中,二者很容易混淆。然而从成因上来看,凝块石主要为沉积作用的直接产物;而豹斑灰岩则属于成岩的范畴,一般多源自次生白云石的产生,其中无机成因可包括构造成因、不均匀白云石化成因以及溶蚀成因等等[18⁃21],而有机成因则多将白云质豹斑归于生物遗迹充填物的白云石化[61⁃62]。事实上昌平组以凝块石为主的微生物岩亦发育了大量被白云石充填的潜穴,如若用有机成因来解释,其和相应的豹斑灰岩相比,仍具有较为明显的微观差异:1)凝块石中的潜穴可见十分清晰的微生物结构和颗粒构造,而豹斑灰岩的潜穴则多由粉—细白云石晶粒构成,基本不具特殊沉积结构/构造。这可能说明沉积作用是凝块石的决定因素,而且白云石化很有可能发生极早,才可将相关结构、构造及时固定,避免后期成岩作用的再次改造;而豹斑灰岩则或多或少被解释为浅埋藏环境下回流渗透白云石化的产物[18⁃21,61⁃62];2)白云质凝块主要由显微凝块构成,且相关SEM图像中可见菌丝和疑似球状原生白云石。其中网状凝块石基质可包含部分白云质成分,而基质中亦可见包含葛万菌、阿哲菌等蓝细菌的显微凝块结构。这可能说明与豹斑灰岩相比,白云质凝块更可能缘自微生物,而并不是潜穴充填物的后期白云石化,加之潜穴之外的显微凝块结构亦包含大量白云石,即白云石局限于微生物及相关物质发育位置,以此区别于白云石局限于潜穴发育位置的豹斑灰岩。所以“微生物白云石化”可能是解释白云质凝块成因的最好模式,即席内微生物可通过新陈代谢活动,如SRB的诱导作用提高胞外微环境的Mg2+饱和度。加之细胞壁及EPS富含负电荷基团,可促进Mg2+克服机械和化学动力学,进入碳酸盐晶格,最终形成白云石[58⁃59]。
-
(1) 北京西郊下苇甸地区下寒武统昌平组底部含燧石角砾白云岩之上发育的一套豹斑灰岩,实为发育在高能环境下的凝块石灰岩。
(2) 该套凝块石灰岩主要发育在昌平组TST和EHST中,巨观上主要表现为生物层,宏观上按形态可分为斑状凝块石、带状凝块石和网状凝块石3种,其分别发育在EHST的潮下带,TST下部的潮间带上部,TST上部的潮间带下部,其成因多与厚微生物席相关,且分别具有不同的微观结构。其中初始沉积组分受控于沉积大/微环境,而三者的互动决定了凝块石灰岩的最终沉积特征。
(3) 该套凝块石灰岩的白云质凝块并不局限于潜穴位置,其内部见大量保存完好微生物结构和疑似球状原生白云石,因此这种白云质凝块很可能与微生物白云石化密切相关,并以此区别于豹斑局限于生物遗迹位置,且其内部充填物为粉细晶白云石的豹斑灰岩。
Characteristics and Genesis of Thrombolites in the Cambrian Changping Formation, Series 2, Beijing
-
摘要:
北京西郊下苇甸地区下寒武统昌平组底部含燧石角砾白云岩之上发育的一套豹斑灰岩,实为发育在高能环境下的凝块石灰岩,宏观形态上可包括斑状凝块石、带状凝块石和网状凝块石3种,其成因多与厚微生物席相关,且分别具有不同的微观结构。露头实测和镜下观察结果表明凝块石的初始沉积组分受控于沉积大/微环境,且三者的互动决定了凝块石的最终沉积特征。该套沉积成因的凝块石,其白云质凝块不局限于潜穴,且潜穴内部见大量保存完好微生物结构和疑似球状原生白云石,因此这种白云质凝块很可能与微生物白云石化密切相关,并以此区别于局限于生物遗迹位置,且内部充填物为粉细晶白云石的成岩成因的豹斑灰岩。 Abstract:Developing above the chert breccia dolomites at the bottom of the Cambrian Changping Formation, Series 2, western Beijing, is a set of so⁃called leopard limestones that are actually thrombolites in a high⁃energy environment and macroscopically include spotted, zonary, and reticular thrombolites. With various microstructures, their geneses are always related to the thick microbial mat. According to the results of outcrop observation and thin section identification, initial fabrics are controlled by the sedimentary and microenvironment in the microbial mat, and their interactions result in the final characteristics of the thrombolites. Thrombolites derived from dolomitic clots of sedimentary origin are not restricted in burrows, where well⁃preserved microfabrics and primary dolomites are found, and are closely related to the microbial dolomitization. These phenomena show the difference between sedimentary thrombolites and diagenetic leopard limestones whose leopard clots are limited to the location of the organic hieroglyph and filled with fine⁃grained dolomites. -
Key words:
- Beijing /
- Cambrian /
- Series 2 /
- thrombolites /
- leopard limestone /
- microbial mat
-

图 2 北京西郊寒武系第二统昌平组生物层与凝块石
(a)巨观生物层,下部岩性为凝块石白云岩,上部为泥质灰岩,具叠层构造,EHST;(b)宏观斑状凝块石(下)与叠层石灰岩(上)共生,EHST;(c)宏观带状凝块石(下)与叠层石砂屑灰岩(上)共生,TST
Figure 2. Biostromes and thrombolites in the Cambrian Changping Formation, Series 2, western Beijing
图 3 斑状凝块石特征
(a)宏观特征;(b)新鲜面;(c)微观特征,镜下见由微晶方解石构成的基质以及由细晶—粉晶白云石构成的显微凝块;(d)第一种显微凝块结构,黄色线条圈起来的为砂屑,红色直线箭头所指为馒头状鲕粒,红色弧线箭头所指为同心圆状鲕粒,橙色直线箭头所指为泥晶—微亮晶胶结物,紫红色直线箭头所指为放射纤维状结壳;(e)红色圈层及红色弧线箭头所指为放射状鲕粒;(f)第二种显微凝块结构,紫色直线箭头指示团块状泥晶;(g)紫色曲线箭头指示绿藻节片;(h)第二种显微凝块结构,红色直线箭头指示砂屑,橙色直线箭头指示生物碎屑
Figure 3. Characteristics of porphyritic thrombolites
图 4 带状凝块石特征
(a)叠层石与带状凝块石伴生;(b)宏观上凝块呈长条状,边缘向上翘起;(c)红色曲线箭头指示核形石,红色直线箭头指示砂屑;(d)蓝色直线箭头处为弥散状泥晶显微叠层结构;(e)红色虚线处为垂直纹层的钻孔结构;(f)基质中第一种显微凝块,以弥散状泥晶组分为主,紫色直线箭头指示泥晶组分,蓝色直线箭头指示显微叠层结构;(g)基质中第二种显微凝块,以团块状泥晶和包壳颗粒为格架,红色直线箭头指示包壳颗粒,橙色直线箭头指示生物碎屑,紫色直线箭头指示泥晶组分;(h)凝块中的显微凝块,以生物碎片为格架,红色曲线箭头处为馒头状鲕粒,橙色直线箭头指示生物碎屑,紫色直线箭头指示泥晶组分
Figure 4. Characteristics of striped thrombolites
图 5 网状凝块石特征
(a)凝块(蓝色虚线箭头)分布在基质(蓝色直线箭头)之间,整体呈网状;(b)不规则孔洞(红色实线圈出);(c)由生物碎屑(橙色直线箭头)、砂屑(红色直线箭头)以及核形石雏形(橙色虚线箭头)组成的小型凝块;(d)管状葛万菌(红色直线箭头),具暗色管壁和亮色管腔,右上角见黄铁矿化葛万菌,右下角见粒状黄铁矿(橙色直线箭头);(e)西兰花状阿哲菌集合体(红色直线箭头),单体呈空心花朵状;(f)窗格状孔隙(红色直线箭头)与由丝状菌组成的网状体;(g)与白云石共生的菌丝(红色直线箭头),SEM图像;(h)球状白云石(红色直线箭头)与粒状白云石(橙色直线箭头),SEM图像
Figure 5. Characteristics of reticular thrombolites
-
[1] Riding R. Classification of microbial carbonates[M]//Riding R. Calcareous algae and stromatolites. Berlin, Heidelberg: Springer, 1991: 21-51. [2] Riding R, Fan J S. Ordovician calcified algae and cyanobacteria, northern Tarim Basin subsurface, China[J]. Palaeontology, 2001, 44(4): 783-810. [3] Riding R. Microbial carbonate abundance compared with fluctuations in metazoan diversity over geological time[J]. Sedimentary Geology, 2006, 185(3/4): 229-238. [4] Riding R. Structure and composition of organic reefs and carbonate mud mounds: Concepts and categories[J]. Earth-Science Reviews, 2002, 58(1/2): 163-231. [5] 梅冥相. 微生物碳酸盐岩分类体系的修订:对灰岩成因结构分类体系的补充[J]. 地学前缘,2007,14(5):222-234. Mei Mingxiang. Revised classification of microbial carbonates: Complementing the classification of limestones[J]. Earth Science Frontiers, 2007, 14(5): 222-234. [6] Aitken J D. Classification and environmental significance of cryptalgal limestones and dolomites, with illustrations from the Cambrian and Ordovician of southwestern Alberta[J]. Journal of Sedimentary Petrology, 1967, 37(4): 1163-1178. [7] Aitken J D, Narbonne G M. Two occurrences of Precambrian thrombolites from the Mackenzie Mountains, northwestern Canada[J]. Palaios, 1989, 4(4): 384-388. [8] Shapiro R S. A comment on the systematic confusion of thrombolites[J]. Palaios, 2000, 15(2): 166-169. [9] Feldmann M, McKenzie J A. Stromatolite-thrombolite associations in a modern environment, Lee Stocking Island, Bahamas[J]. Palaios, 1998, 13(2): 201-212. [10] Planavsky N, Ginsburg R N. Taphonomy of modern marine Bahamian microbialites[J]. Palaios, 2009, 24(1/2): 5-17. [11] Dupraz C, Reid R P, Braissant O, et al. Processes of carbonate precipitation in modern microbial mats[J]. Earth-Science Reviews, 2009, 96(3): 141-162. [12] Tang D J, Shi X Y, Jiang G Q, et al. Environment controls on Mesoproterozoic thrombolite morphogenesis: A case study from the North China Platform[J]. Journal of Palaeogeography, 2013, 2(3): 275-296. [13] Tang D J, Shi X Y, Jiang G Q, et al. Microfabrics in Mesoproterozoic microdigitate stromatolites: Evidence of biogenicity and organomineralization at micron and nanometer scales[J]. Palaios, 2013, 28(3): 178-194. [14] Arp G, Helms G, Karlinska K, et al. Photosynthesis versus exopolymer degradation in the formation of microbialites on the Atoll of Kiritimati, Republic of Kiribati, Central Pacific[J]. Geomicrobiology Journal, 2012, 29(1): 29-65. [15] 梅冥相,Latif K,刘丽,等. 光合作用生物膜建造的凝块:来自于辽东半岛芙蓉统长山组凝块石生物丘中的一些证据[J]. 古地理学报,2019,21(2):254-277. Mei Mingxiang, Latif K, Liu Li, et al. Clots built by photosynthetic biofilms: Evidences from thrombolite bieherms of the Changshan Formation of Cambrian Furongian in Liaodong Peninsula[J]. Journal of Palaeogeography (Chinese Edition), 2019, 21(2): 254-277. [16] 梅冥相. 微生物席的特征和属性:微生物席沉积学的理论基础[J]. 古地理学报,2014,16(3):285-304. Mei Mingxiang. Feature and nature of microbial-mat: Theoretical basis of microbial-mat sedimentology[J]. Journal of Palaeogeography (Chinese Edition), 2014, 16(3): 285-304. [17] 梅冥相. 微生物席沉积学:一个年轻的沉积学分支[J]. 地球科学进展,2011,26(6):586-597. Mei Mingxiang. Microbial-mat sedimentology: A young branch from sedimentology[J]. Advances in Earth Science, 2011, 26(6): 586-597. [18] 陈云峰,吴淦国,王根厚. 北京周口店豹皮灰岩的变形特征[J]. 地质通报,2007,26(6):769-775. Chen Yunfeng, Wu Ganguo, Wang Genhou. Deformation characteristics of leopard limestone in Zhoukoudian, Beijing, China[J]. Geological Bulletin of China, 2007, 26(6): 769-775. [19] 梅冥相. 华北寒武系二级海侵背景下的沉积趋势及层序地层序列:以北京西郊下苇甸剖面为例[J]. 中国地质,2011,38(2):317-337. Mei Mingxiang. Depositional trends and sequence-stratigraphic successions under the Cambrian second-order transgressive setting in the North China Platform: A case study of the Xiaweidian section in the western suburb of Beijing[J]. Geology in China, 2011, 38(2): 317-337. [20] 龙刚,黄萍,林剑怀,等. 徐州地区寒武系豹皮灰岩的岩性特征及其成因机制分析[J]. 地质学刊,2013,37(1):67-70. Long Gang, Huang Ping, Lin Jianhuai, et al. Lithologic characteristics of leopard limestone in Cambrian Period and analysis of its formation mechanism in Xuzhou northern [J]. Journal of Geology, 2013, 37(1): 67-70. [21] 李波,颜佳新,薛武强,等. 四川广元地区中二叠世斑状白云岩成因及地质意义[J]. 地球科学:中国地质大学学报,2012,37(增刊2):136-146. Li Bo, Yan Jiaxin, Xue Wuqiang, et al. Origin of patchy dolomite and its geological signification from Middle Permian, Guangyuan, Sichuan province[J]. Earth Science:Journal of China University of Geosciences, 2012, 37(Suppl. 2): 136-146. [22] Rowland S M, Shapiro R S. Reef patterns and environmental influences in the Cambrian and earliest Ordovician[M]//Kiessling W, Flügel E, Golonka J. Phanerozoic reef patterns. Tulsa: SEPM Special Publication, 2002: 95-128. [23] Feng Z Z, Chen J X, Wu S H. Lithofacies palaeogeography of Early Paleozoic of North China Platform[J]. Acta Sedimentologica Sinica, 1989, 7(4): 16-55. [24] 白莹,罗平,刘伟,等. 北京西郊丁家滩剖面寒武系第二统昌平组核形石特征及成因[J]. 现代地质,2019,33(3):587-597. Bai Ying, Luo Ping, Liu Wei, et al. Characteristics and origin of oncolite from Changping Formation in the Series 2 of Cambrian in western Beijing[J]. Geoscience, 2019, 33(3): 587-597. [25] 梅冥相,胡媛,孟庆芬. 大连金州湾寒武系毛庄组微生物碳酸盐岩生物丘复合体[J]. 地质学报, 2020, 94(2): 375-395. Mei Mingxiang, Hu Yuan, Meng Qingfen. Bioherm complex madding up of microbial carbonates in the Cambrian Maozhuang Formation, Jinzhouwan section in the Dalian city of Liaoning province in northeastern China[J]. Acta Geologica Sinica, 94(2): 375-395. [26] 杨孝群,李忠. 微生物碳酸盐岩沉积学研究进展:基于第33届国际沉积学会议的综述[J]. 沉积学报,2018,36(4):639-650. Yang Xiaoqun, Li Zhong. Research progress in sedimentology of microbial carbonate rocks: A review based on the 33rd International Sedimentological Congress[J]. Acta Sedimentologica Sinica, 2018, 36(4): 639-650. [27] 韩作振,陈吉涛,迟乃杰,等. 微生物碳酸盐岩研究:回顾与展望[J]. 海洋与第四纪地质,2009,29(4):29-38. Han Zuozhen, Chen Jitao, Chi Naijie, et al. Microbial carbonates: A review and perspectives[J]. Marine Geology & Quaternary Geology, 2009, 29(4): 29-38. [28] Tucker M, Perri E, Slowakiewicz M, et al. Mineral precipitates in modern microbial mats: Crystallites, spheroids, bacteria and viruses[C]//Proceedings of the 33rd IAS international meeting of sedimentology. Toulouse: IMS, 2017. [29] Pace A, Bourillot R, Bouton A, et al. Initiation of Stromatolite Formation at the interface of oxygenic-anoxygenic phoyosynthesis[C]//Proceedings of the 33rd IAS international meeting of sedimentology. Toulouse: IMS, 2017. [30] Merz-Preiß M. Calcification in cyanobacteria[M]//Riding R E, Awramik S M. Microbial sediments. Berlin, Heidelberg: Springer, 2000: 50-56. [31] Pentecost A, Riding R. Calcification in cyanobacteria[M]//Leadbeater B S C, Riding R. Biomineralization of Lower plants and animals. Oxford: Oxford University Press, 1986: 73-90. [32] Thompson J B, Ferris F G. Cyanobacterial precipitation of gypsum, calcite, and magnesite from natural alkaline lake water[J]. Geology, 1990, 18(10): 995-998. [33] Merz M U E. The biology of carbonate precipitation by cyanobacteria[J]. Facies, 1992, 26(1): 81-101. [34] Arp G, Reimer A, Reitner J. Photosynthesis-induced biofilm calcification and calcium concentrations in Phanerozoic oceans[J]. Science, 2001, 292(5522): 1701-1704. [35] Riding R. An atmospheric stimulus for cyanobacterial-bioinduced calcification ca. 350 million years ago?[J]. Palaios, 2009, 24(10): 685-696. [36] 项礼文, 朱兆玲, 李善姬, 等. 中国地层典(寒武系) [M]. 北京:地质出版社,2000:1-95. Xiang Liwen, Zhu Zhaoling, Li Shanji, et al. Stratigraphical Lexicon of China: Cambrian[M]. Beijing: Geological Publishing House, 2000: 1-95. [37] 卢衍豪.中国的寒武系[M]. 北京:科学出版社,1962:1-119. Lu Yanhao. Cambrian in China[M]. Beijing: Science Press, 1962: 1-119. [38] 卢衍豪, 朱兆玲, 钱义元, 等. 中国寒武纪地层对比表及说明书[C]//中国各纪地层对比表及说明书. 北京:科学出版社, 1982:28-54. [ Lu Yanhao, Zhu Zhaoling, Qian Yiyuan, et al. Stratigraphical correlation tables and their directions for the Cambrian in China[C]// Stratigraphical Correlation Tables and Their Directions for Each Periods. Beijing: Science Press, 1982:28-54.] [39] 史晓颖,陈建强,梅仕龙. 华北地台东部寒武系层序地层年代格架[J]. 地学前缘,1997,4(3/4):161-173. Shi Xiaoying, Chen Jianqiang, Mei Shilong. Cambrian sequence chronostratigraphic frame work of the North China Platform[J]. Earth Science Frontiers, 1997, 4(3/4): 161-173. [40] Peng S C. The newly-developed Cambrian biostratigraphic succession and chronostratigraphic scheme for South China[J]. Chinese Science Bulletin, 2009, 54(22): 4161-4170. [41] Reid R P, Macintyre I G. Microboring versus recrystallization: Further insight into the micritization process[J]. Journal of Sedimentary Research, 2000, 70(1): 24-28. [42] Pratt B R. Calcification of cyanobacterial filaments: Girvanella and the origin of Lower Paleozoic lime mud[J]. Geology, 2001, 29(9): 763-766. [43] Perry C T. Grain susceptibility to the effects of microboring: Implications for the preservation of skeletal carbonates[J]. Sedimentology, 1998, 45(1): 39-51. [44] Golubic S, Campbell S E, Drobne K, et al. Microbial endoliths: A benthic overprint in the sedimentary record, and a paleobathymetric cross-reference with foraminifera[J]. Journal of Paleontology, 1984, 58: 351-361. [45] Garcia-Pichel F. Plausible mechanisms for the boring on carbonates by microbial phototrophs[J]. Sedimentary Geology, 2006, 185(3/4): 205-213. [46] Gerdes G, Dunajtschik-Piewak K, Riege H, et al. Structural diversity of biogenic carbonate particles in microbial mats. Sedimentology, 1994, 41(6): 1273-1294. [47] 梅冥相,刘丽,胡媛. 北京西郊寒武系凤山组叠层石生物层[J]. 地质学报,2015,89(2):440-460. Mei Mingxiang, Liu Li, Hu Yuan. Stromatolitic biostrome of the Cambrian Fengshan Formation at the Xiaweidian section in the western suburb of Beijing, North China[J]. Acta Geologica Sinica, 2015, 89(2): 440-460. [48] Mobberley J M, Khodadad C L M, Foster J S. Metabolic potential of lithifying cyanobacteria-dominated thrombolitic mats[J]. Photosynthesis Research, 2013, 118(1/2): 125-140. [49] Pinckney J L, Reid R P. Productivity and community composition of stromatolitic microbial mats in the Exuma Cays, Bahamas[J]. Facies, 1997, 36: 204-207. [50] 刘闯,钟建华,曹梦春,等. 泥裂影响因素及其研究意义综述[J]. 地球科学与环境学报,2017,39(4):539-550. Liu Chuang, Zhong Jianhua, Cao Mengchun, et al. Review on influence factors of mud cracks and their significance[J]. Journal of Earth Sciences and Environment, 2017, 39(4): 539-550. [51] 贾进华. 古海岸带碎屑潮汐环境沉积微相与砂体分布:以塔中地区志留系为例[J]. 中国矿业大学学报,2019,48(1):110-123. Jia Jinhua. Sedimentary microfacies and sandbody distribution in the clastic tidal environment of the ancient coastal zone: A case study of Silurian in Tazhong area, Tarim Basin[J]. Journal of China University of Mining & Technology, 2019, 48(1): 110-123. [52] Riding R. Cyanobacterial calcification, carbon dioxide concentrating mechanisms, and Proterozoic-Cambrian changes in atmospheric composition[J]. Geobiology, 2006, 4(4): 299-316. [53] 刘丽静,杨海军,潘文庆,等. 中国新疆塔里木盆地上奥陶统良里塔格组的钙藻化石[J]. 微体古生物学报,2012,29(1):18-38. Liu Lijing, Yang Haijun, Pan Wenqing, et al. Calcareous algae from the Upper Ordovician Lianglitage Formation in the Tarim Basin, Xinjiang, China[J]. Acta Micropalaeontologica Sinica, 2012, 29(1): 18-38. [54] 王龙,Latif K,Riaz M,等. 微生物碳酸盐岩的成因、分类以及问题与展望:来自华北地台寒武系微生物碳酸盐岩研究的启示[J]. 地球科学进展,2018,33(10):1005-1023. Wang Long, Latif K, Riaz M, et al. The genesis, classification, problems and prospects of microbial carbonates: Implications from the Cambrian carbonate of North China Platform[J]. Advances in Earth Science, 2018, 33(10): 1005-1023. [55] Sánchez-Román M, McKenzie J A, De Luca Rebello Wagener A, et al. Presence of sulfate does not inhibit low-temperature dolomite precipitation[J]. Earth and Planetary Science Letters, 2009, 285(1/2): 131-139. [56] Louyakis A S, Mobberley J M, Vitek B E, et al. A study of the microbial spatial heterogeneity of Bahamian thrombolites using molecular, biochemical, and stable isotope analyses[J]. Astrobiology, 2017, 17(5): 413-430. [57] Louyakis A S, Gourlè H, Casaburi G, et al. A year in the life of a thrombolite: Comparative metatranscriptomics reveals dynamic metabolic changes over diel and seasonal cycles[J]. Environmental Microbiology, 2018, 20(2): 842-861. [58] 由雪莲,孙枢,朱井泉,等. 微生物白云岩模式研究进展[J]. 地学前缘,2011,18(4):52-64. You Xuelian, Sun Shu, Zhu Jingquan, et al. Progress in the study of microbial dolomite model[J]. Earth Science Frontiers, 2011, 18(4): 52-64. [59] 许杨阳,刘邓,于娜,等. 微生物(有机)白云石成因模式研究进展与思考[J]. 地球科学,2018,43(增刊1):63-70. Xu Yangyang, Liu Deng, Yu Na, et al. Advance and review on microbial/organogenic dolomite model[J]. Earth Science, 2018, 43(Suppl. 1): 63-70. [60] 肖恩照, 王皓, 覃英伦, 等. 寒武纪芙蓉统均一石沉积组构及环境特征:以河北涞源长山组为例[J]. 沉积学报, 2020, 38(1): 76-90. Xiao Enzhao, Wang Hao, Qin Yinglun, et al. Sedimentary fabrics and environmental characteristics of leiolite in Cambrian: A case study from the Changshan Formation in Laiyuan city, Hebei provice[J]. Acta Sedimentologica Sinica, 2020, 38(1): 76-90. [61] 刘梦瑶,齐永安,史云鹤,等. 华北寒武纪—奥陶纪豹皮状碳酸盐岩系生物扰动成因[J]. 沉积学报,2020,38(1):91-103. Liu Mengyao, Qi Yong’an, Shi Yunhe, et al. Formation mechanism of Cambrian-Ordovician bioturbated dolomites in North China[J]. Acta Sedimentologica Sinica, 2020, 38(1): 91-103. [62] 董小波,牛永斌. 豫西北奥陶系马家沟组三段豹斑灰岩的生物潜穴成因及成岩演化[J]. 现代地质,2015,29(4):833-843. Dong Xiaobo, Niu Yongbin. Biological burrow explanation of leopard limestone and its diagenetic evolution in the third member of Majiagou Formation in Ordovician, northwest of Henan province[J]. Geoscience, 2015, 29(4): 883-843. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1174
- HTML全文浏览量: 161
- PDF下载量: 111
- 被引次数: 0

