



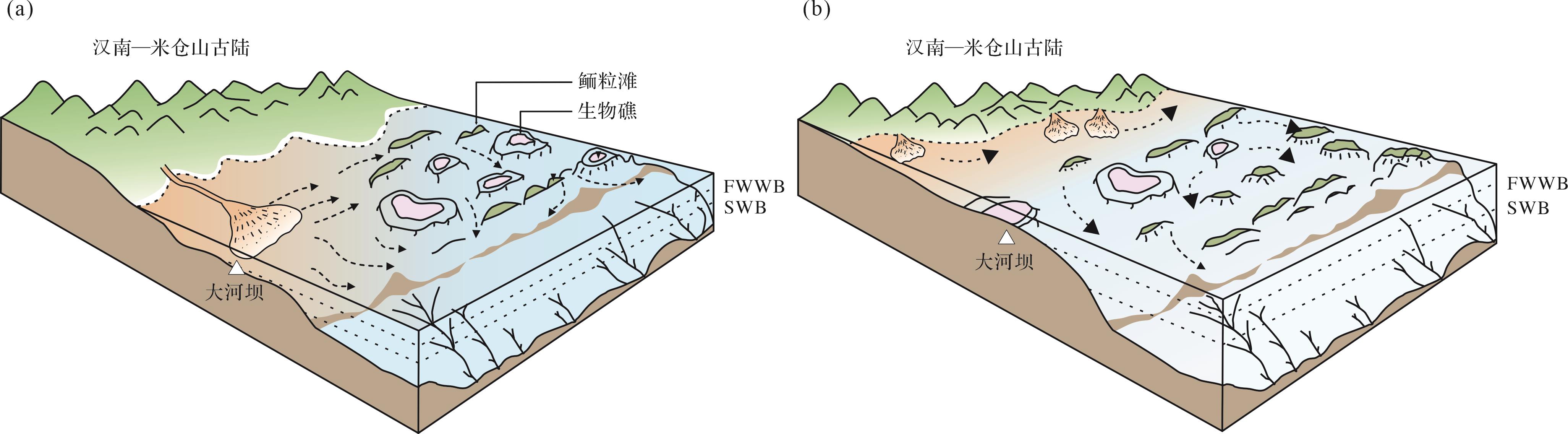

-
生物礁作为海洋高等级生态系统的重要组成部分,是研究现代和地史时期生态与环境协同演化的良好载体。生物礁主要由原地生物参与构建形成的具有隆起地形的特殊碳酸盐岩体[1⁃2],也是重要的碳酸盐岩型油气资源储库[3⁃4]。借鉴于现代珊瑚礁生长模式解读地质记录中生物礁发育过程时,前人认为生物礁的首要特征在于造礁生物具有坚固钙质骨骼,其建造者既包括营固着生长的多细胞真核生物,如底栖钙藻类、古杯、海绵、珊瑚等,也包括能够诱导或影响碳酸盐矿物沉淀的微生物群落[1,5]。此外,早期胶结作用也可促进造礁生物和其他共生颗粒类型的固定[1,6⁃8]。
前寒武纪以微生物主导的具纹层状和凝块状结构的礁体为主[9],在前寒武至寒武纪之交时造礁分子、类型和规模均发生了明显变化[10]。埃迪卡拉纪晚期(~580-541 Ma)出现了后生动物如Cloudina等与微生物共同形成礁灰岩的现象[11⁃12],但Cloudina是否确实具有造礁能力仍然存在争议[13⁃14]。寒武纪早期古杯动物在全球范围内广泛发育,其既可以独立形成礁格架,也可参与微生物主导的礁体结构构建[15⁃21]。在寒武纪第二世和苗岭世之交的古杯动物灭绝事件之后,微生物主导的生物礁建造再次出现[22],且后生动物(如海绵等)参与程度较低,这种情况一直持续到奥陶纪生物大辐射事件之初[23]。
寒武纪早期古杯动物与钙质微生物在生物礁建造过程中的贡献和生态关系值得探索[16,24]。古杯动物造礁最早出现在西伯利亚纽芬兰世第二期(Tommotian期),在第二世达到演化顶峰后快速消亡[15,25⁃26]。华南板块西北缘汉南—米仓山地区寒武系第三阶仙女洞组底部产出目前已知国内最早的古杯动物化石[27],系统古生物学工作厘定了古杯动物的时代以及区域分布[28⁃29]。前人大量工作显示古杯化石集中出现在礁灰岩层位[30⁃36];以古杯动物和钙质微生物共同参与形成的生物礁常呈缓丘状、透镜状,或高大隆起状,围绕汉南—米仓山古陆呈环带状分布[37⁃39]。仙女洞组礁体特殊性在于其陆源碎屑组分含量高,古杯—钙质微生物礁如何应对浑浊水体及泥沙海底环境目前还不明确。因此,了解寒武纪早期浅水碳酸盐组分与陆源碎屑组分混合沉积背景下生物礁发育机理和浅水碳酸盐工厂建造过程是一个值得探索的科学问题。本文综合分析了陕南勉县大河坝一带仙女洞组古杯—钙质微生物礁的岩相学特征,着重阐述了浅海混积体系背景下早期后生动物与微生物礁建造的沉积学过程,为古环境学、沉积学和储层地质学相关研究提供了一个参考实例。
-
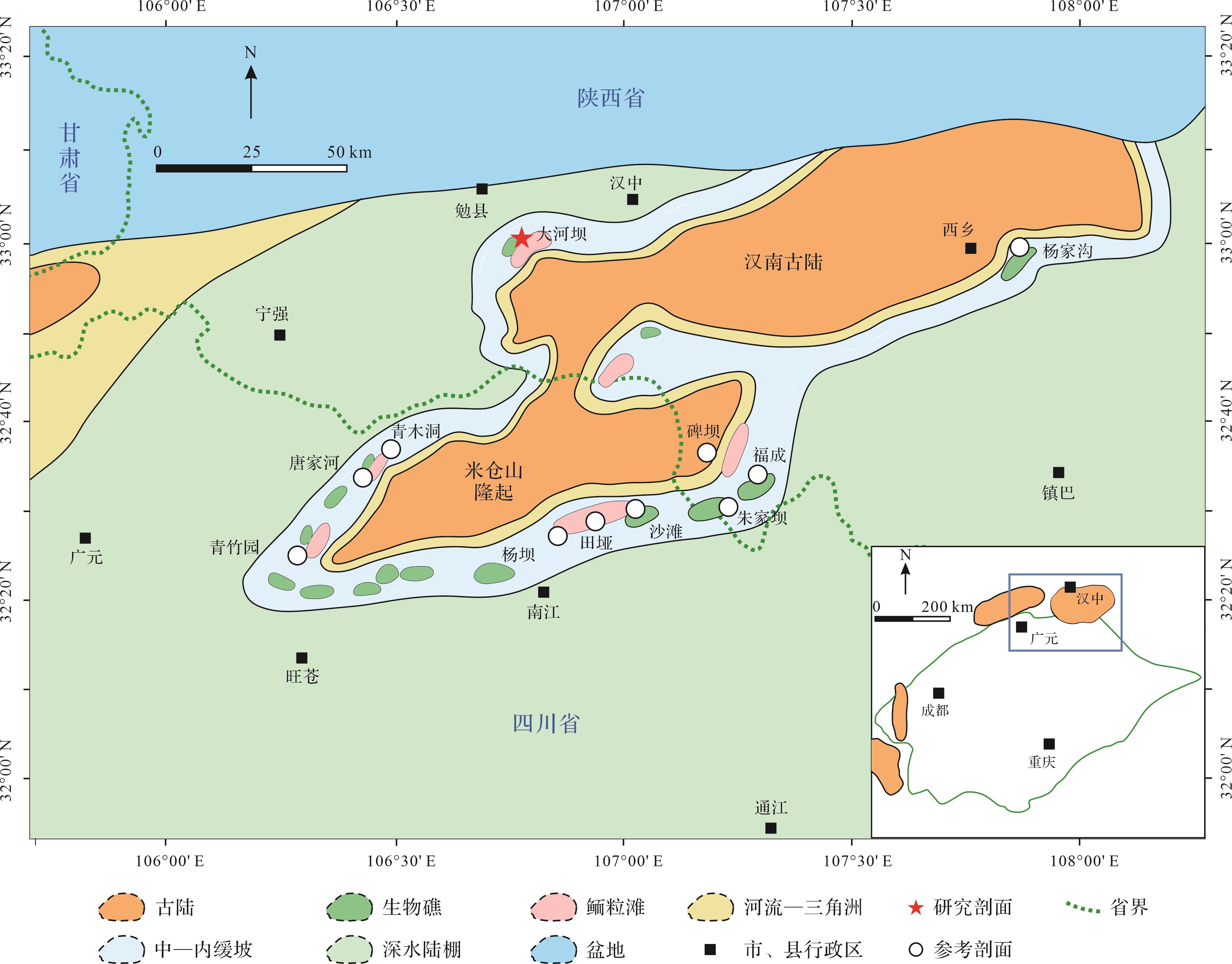
汉南—米仓山地区位于上扬子板块北缘与秦岭造山带之间过渡地带,以新元古代经历一系列构造活动后拼合的汉南古陆和米仓山隆起带为中心[40](图1),在大地构造上西临龙门山构造带,东接大巴山褶皱带,北与勉略缝合带相接,南邻四川盆地[36]。寒武纪第二期与第三期之交研究区因大规模海侵[41]而以深水陆棚环境为主,在第三期海退过程中古陆面积逐步扩大,周缘以碳酸盐与碎屑组分的混合沉积为主[36],由陆向海依次发育河流—三角洲、混积台地(滨岸—过渡带)和深水陆棚(图1),至第三期与第四期之交转为三角洲沉积为主[30]。

图 1 研究区位置及寒武纪第三期古地理图
Figure 1. Location and paleogeography (Cambrian Age 3 ) of the study area
研究区内寒武系沉积序列存在大规模的缺失,主要发育第二统(图2)。由下至上,郭家坝组岩性为钙质泥岩、粉砂质泥岩和泥质粉砂岩[43];仙女洞组为砂质(含砂)鲕粒岩、微生物岩、含古杯化石灰岩,间歇性发育钙质砂岩和粉砂岩等[35,38];阎王碥组底部由含钙粉砂岩构成的红层,向上为灰色粉砂岩、砂岩和砾岩等[39,43]。本研究的陕西勉县阜川镇大河坝剖面(GPS:32°56′39″ N,106°41′45″ E)大部分露头位于上山公路旁,地层出露良好,接触关系清晰,厚度66.7 m。古杯—钙质微生物礁保存于仙女洞组中上部。
-
剖面底部为灰褐色中薄层纹层状含钙粉砂岩(图3a),以及泥质粉砂岩与粉砂质泥岩互层;水平层理发育,含较多生物潜穴(图3b)。向上为厚11.62 m的浅灰色—灰褐色厚层含砾岩屑中—粗砂岩(图3c,d),颗粒以岩屑和石英为主,分选和磨圆极差。接近生物礁发育层位以中薄层钙质粉砂岩为主,厚2.65 m(图3e)。生物礁宏观上呈穹窿状,宽5.7 m,高3.1 m(图3f)。生物礁之上为灰色中层状含钙岩屑砂岩,平行层理发育,厚11.39 m。近仙女洞组顶部为厚度约3 m的灰色厚层含砂亮晶鲕粒灰岩,与上覆阎王碥组底部红色薄层状泥质粉砂岩整合接触。

图 3 大河坝剖面仙女洞组露头
Figure 3. Photographs showing the outcrops characteristics of Xiannüdong Formation at the Daheba section
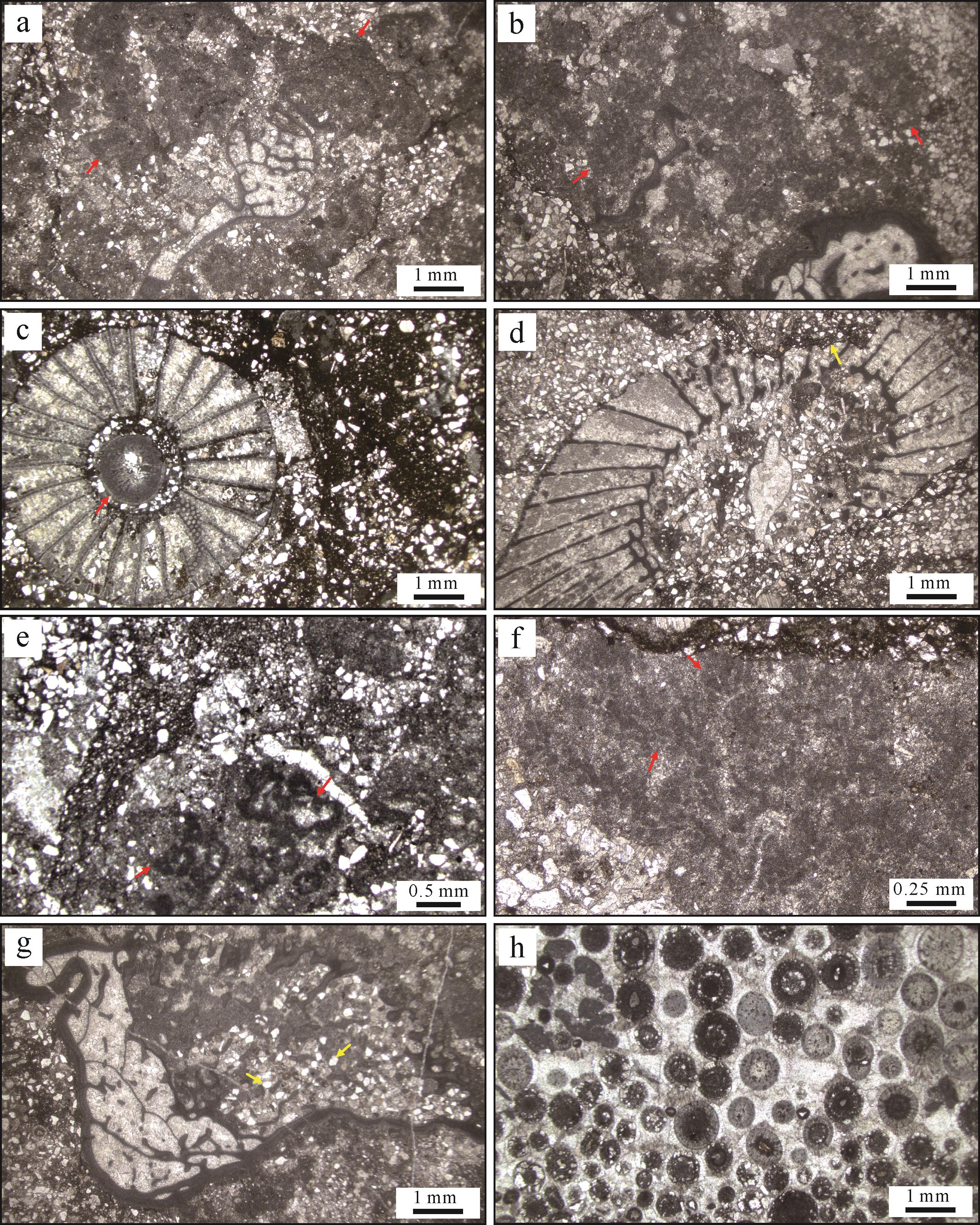
含砂古杯—凝块灰岩构成的主礁体边界清晰,具有向上隆起外观(图3f),主礁体两侧均见高度不足2 m的小礁体,礁间为鲕粒灰岩(图3f)。生物礁从底部至近顶部可依次分为凝块石主导、古杯动物主导,以及古杯—凝块石共生三个阶段(图3f)。微生物凝块结构呈凝絮麻粒状,一般不超过1 cm(图4)。古杯动物主要集中在礁体中部(图3g),杯状外形,长1~4 cm,直径多小于2 cm,大多保存完整,少量为古杯碎片,整体呈层状分布(图4a,b)。在礁体中部至上部可见古杯动物与微生物凝块的共生关系(图4a,b),且向上逐趋明显(图4c,d)。礁间鲕粒密集(图3h)。

图 4 大河坝仙女洞组造礁古杯动物和钙质微生物抛光面特征
Figure 4. Polished slabs showing archaeocyaths and calcified microbes identified in the Xiannüdong Formation of the Daheba section
不规则古杯类常见隐晶质外壁和曲板,截面直径一般在0.2~1 cm,整体保存状态较好(图5a,b)。规则古杯类杯口直径常在0.3~1 cm,具有隐晶质外壁和隔板,少量可见横板(图5c,d)。古杯动物呈单体产出,也可与钙质微生物共生(图5a,b),在生物礁发育阶段Ⅱ和Ⅲ密集产出。大量陆源黏土至粉砂级组分常与古杯动物相邻出现,或者直接分布于古杯隔板间或中空结构(图5c),也可见陆源粉砂磨蚀古杯外壁特征(图5d)。

图 5 大河坝生物礁层的岩相学特征
Figure 5. Lithofacies of photomicrographs showing the petrological characteristics of the Daheba reef
钙质微生物主要类型为附枝菌和肾形菌。肾形菌一般为次圆形房室状,单个菌体大小在200~500 µm,内部具中空结构,周围含细粉砂和黏土(图5e)。附枝菌单个分枝宽度一般在100~200 µm,单体纵切面分叉呈树枝状,横切面呈泥晶粒状(图5f),纵切面呈灌木丛状,亮晶方解石胶结,或直接被陆源碎屑组分包裹(图4c)。生物礁内钙质微生物主要与古杯动物共生,以黏结或包绕方式为主(图4);少量单独产出,形成簇状或小型树枝状结构(图4c)。
鲕粒主要分布于礁间,与少量古杯、三叶虫等化石碎片伴生,其原生结构主要为放射状或同心—放射状,部分鲕粒内部受成岩改造保存为泥晶结构(图5h);还可见部分鲕粒内部原生结构被粗粉晶—细晶(50~200 µm)白云石交代。鲕粒间主要为未受到白云石化作用影响的亮晶方解石胶结物,具有期次性(图5h)。
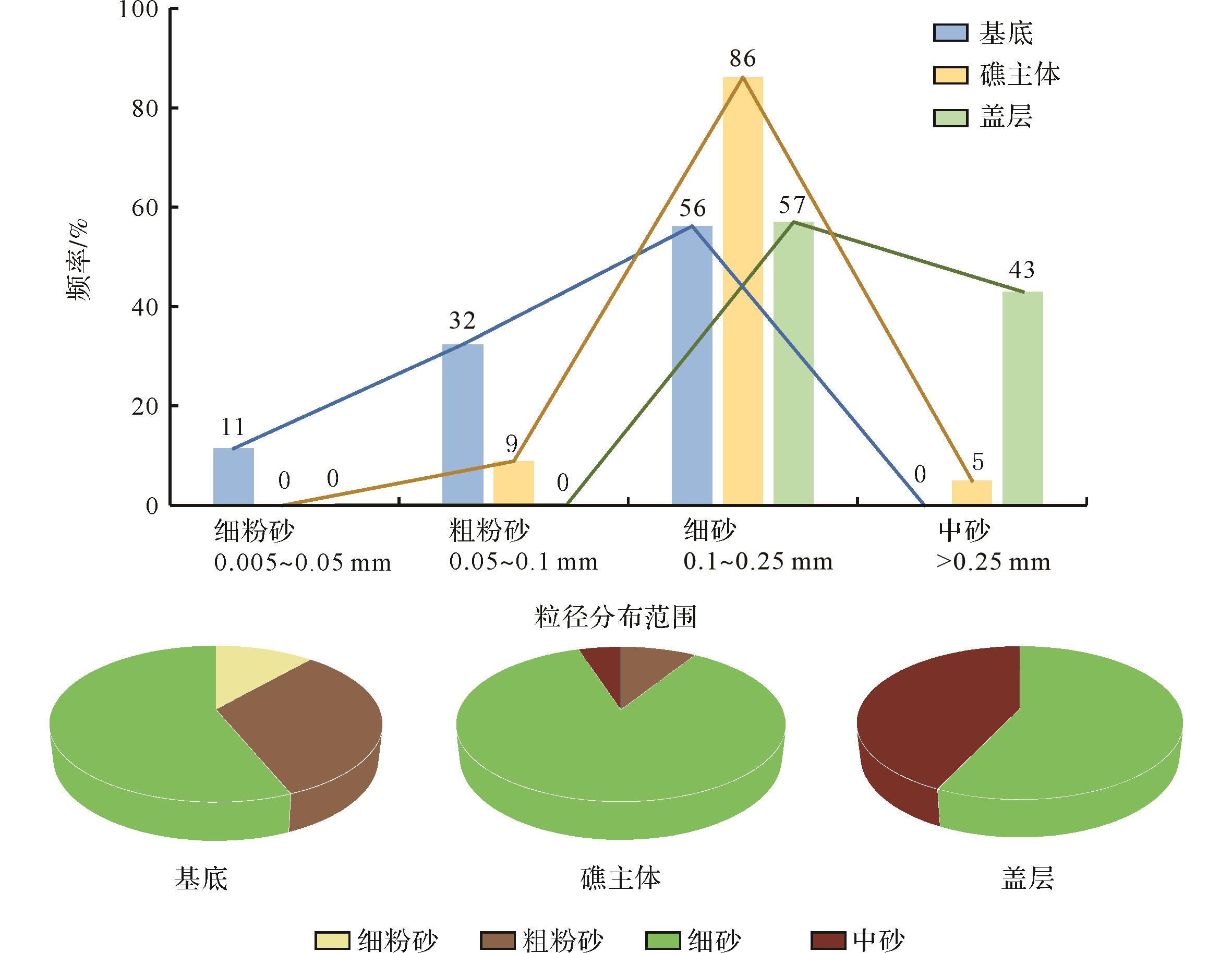
生物礁内混入的陆源碎屑包括黏土、粉砂和细砂级组分,颗粒类型主要为石英和岩屑,少量为长石(不足5%)。这些陆源碎屑组分多分布在古杯动物和微生物组构的相邻层(图4),部分存在于古杯体腔和钙质微生物周缘(图5a~g)。生物礁内部石英颗粒含量占陆源颗粒总量约30%,分选良好,磨圆度差;岩屑颗粒含量占50%以上,多为次棱角状。其中生物礁基底陆源碎屑颗粒以粗粉砂和细砂为主,礁主体以细砂为主,盖层以细砂和中砂为主(图6)。

图 6 生物礁内部陆源碎屑颗粒粒级及分布
Figure 6. Size distribution of terrigenous particles in the Daheba reef
大河坝剖面岩相类型可分为7种,详细特征见表1。
表 1 大河坝剖面主要岩相类型和特征
Table 1. Lithofacies types and characteristics of the Daheba section
编号 主要特征 沉积构造 沉积环境 LF1 薄层粉砂质泥岩为主,少量为薄层泥岩夹粉砂岩。可见生物活动钻孔,几乎不含生物化石,有机质含量较高,可见沥青充填孔隙 水平层理、见生物钻孔 深水陆棚 LF2 薄层含钙粉砂岩为主,部分为含钙粉砂岩与泥岩/粉砂质泥岩互层,偶夹钙质细砂岩纹层。生物钻孔孔径一般为2 mm,常被钙质粗粉砂—细砂充填 水平层理、生物钻孔非常发育 过渡带 LF3 中薄层钙质岩屑细砂岩与钙质粉砂岩互层,部分为细砂岩夹粉砂岩。钙质组分含量较高,最高可达30%,多以胶结物的形式产出。见海绿石,呈次圆状,磨圆良好,偶见异地搬运鲕粒以及三叶虫化石碎片。陆源碎屑颗粒分选较好,磨圆中等—差;岩屑含量多于石英,次棱角状—次圆状 平行层理 下临滨 LF4 底部和顶部为中厚层含钙石英岩屑中砂岩,分选中等,颗粒之间呈点—线接触;钙质组分以胶结物形式充填陆源颗粒之间孔隙,含量一般不超过10%,且不含生物化石。中部为块状含砾岩屑粗砂岩,分选、磨圆极差,成分成熟度与结构成熟度低,并显示出杂乱堆积的特征。砾石粒径在2~5 mm,砾石间充填细砂至粗砂颗粒以及少量的泥,不含生物碎屑 递变层理、砾石叠瓦构造 洪积扇 LF5 块状含砂(砂质)生物灰岩和凝块石。其中含砂生物灰岩中陆源碎屑组分约为10%~20%,以岩屑为主,石英次之,颗粒分选中等,磨圆较差,多分布于古杯和钙质微生物周缘。古杯常与钙质微生物共生。钙质微生物类型为附枝菌和肾形菌,生物格架之间多为亮晶方解石胶结物。砂质凝块石中陆源碎屑组分含量超过30%,颗粒粒径可达粗粉砂级,分选较好,磨圆差,石英和岩屑颗粒多为次棱角状。古杯含量少。钙质微生物特征受高通量陆源物质混入影响,保存差 未见 生物礁 LF6 厚层含砂鲕粒灰岩。鲕粒粒径一般在0.6~1 mm,同心—放射状结构为主,部分被白云石晶粒交代;亮晶方解石胶结,分选好,可见少量古杯和三叶虫化石碎片。陆源碎屑组分主要为中砂和细砂级石英和岩屑颗粒,分选较好,磨圆中等—较差,颗粒含量占总体约10%~25% 未见 鲕粒滩 LF7 中层状含钙细砂岩和中—细砂岩。陆源碎屑组分分选良好,磨圆中等,颗粒类型包括岩屑(含量约40%)、石英(约50%)、长石(<10%)。颗粒之间呈凹凸接触,多为棱角—次棱角状。钙质组分含量不超过15%,主要为陆源碎屑颗粒胶结物形式,部分云化,未见碳酸盐颗粒 平行层理 上临滨 -
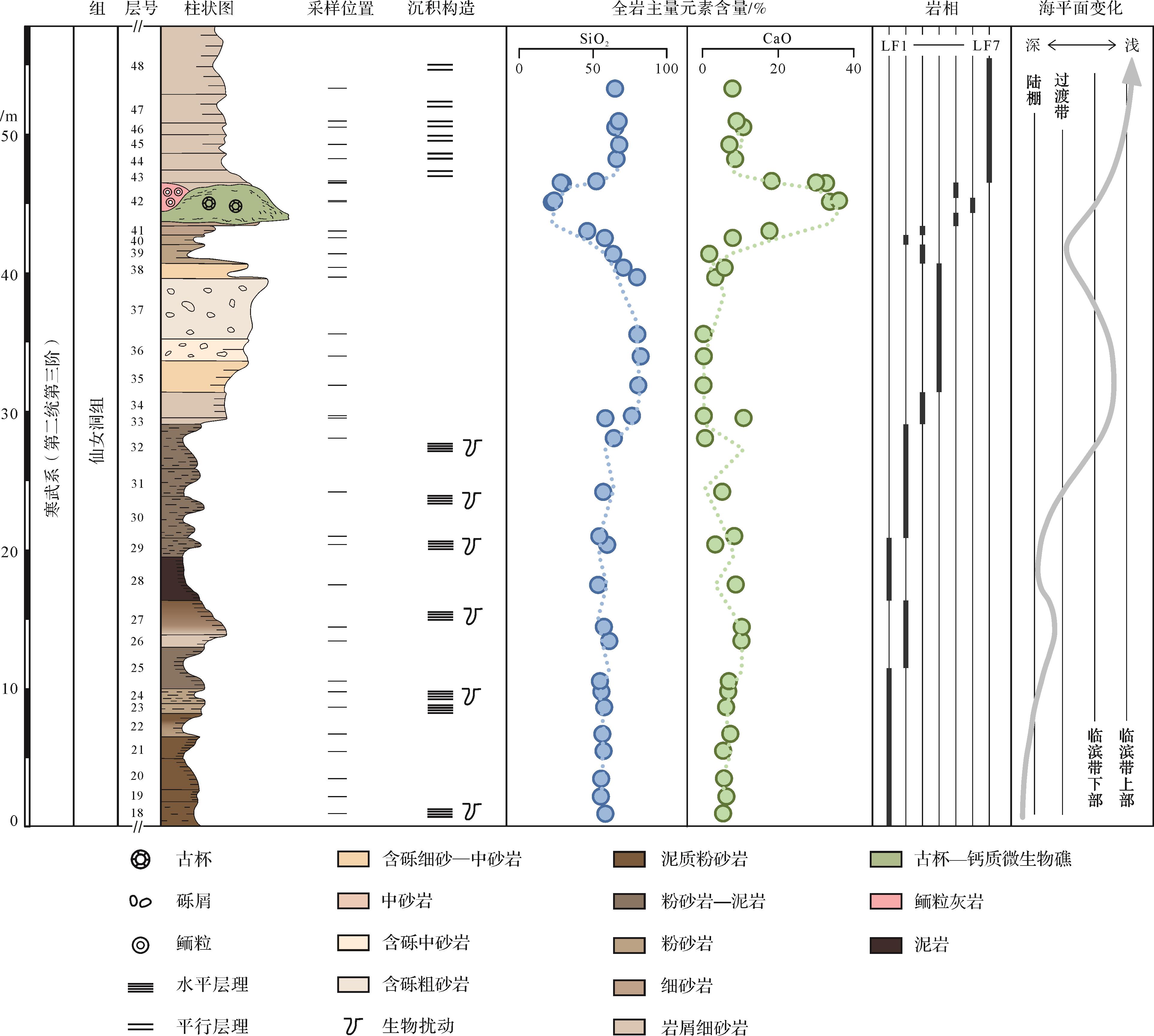
大河坝剖面仙女洞组沉积期环境经历了从滨外带过渡到临滨带下部,经历洪水事件后逐渐转变为临滨带上部的过程,整体水深逐渐变浅(表1、图7)。

图 7 大河坝剖面仙女洞组综合柱状图
Figure 7. Integrated characteristics of lithofacies, sea level, and major element contents at the Daheba section
洪积扇的垫高效应对于形成适合生物礁发育的环境具有一定促进作用。汉南—米仓山古陆南缘和东缘仙女洞组中下部发育的生物礁主要位于混积台地,以钙质细砂岩为基底,形成于潮下带水体能量较强的环境[35⁃38]。位于古陆西侧的研究区仙女洞组底部沉积环境上属于陆棚和滨岸之间过渡带(图3a,b、图7),仙女洞组中下部岩相特征(LF4,表1)指示突发洪水事件,保存约10 m厚粗粒陆源碎屑物质,该过程抬高了沉积底形(图8a)。此后虽经历短暂海侵,但整体海退过程仍促使沉积环境从滨外带快速过渡至临滨带下部,在此背景下生物礁开始发育。

大河坝生物礁的发育与当地陆源输入大幅减少存在关联。滨外环境以粉砂和泥质碎屑组分为主,而滨岸带多中、细砂陆源颗粒(图7),钙质组分主要以胶结物形式存在。生物礁层位中陆源碎屑含量减少且颗粒类型以生物、微生物和鲕粒为主(图5)。元素地球化学证据显示陆源组分输入量指标(SiO2)在生物礁和邻近层位有显著降低,且指示碳酸盐组分含量指标(CaO)在对应层位达到峰值(图7)。这些证据表明古杯—钙质微生物礁形成于陆源输入量显著降低的环境。岩相学特征显示较高的陆源碎屑含量间歇分布于古杯—钙质微生物层间,或古杯格架之间的缝隙;不仅古杯体腔内含砂级石英颗粒,钙质微生物结构内部也含有粉砂—黏土级碎屑组分(图4,5)。相关特征指示古杯—钙质微生物礁建造过程中频繁地受到陆源碎屑组分侵入影响,表现为弱混浊环境对生物礁发育影响有限;阵发性高通量泥沙输入可能暂时中止生物礁的发育。
生物礁下部含砂凝块石的发育(阶段Ⅰ)为后续生物礁的建造提供了硬质基底条件(图3f)。大量古杯和钙质微生物的繁盛出现在礁核中上部(图5),而共存细砂级为主的陆源碎屑组分不断增加且粒径逐渐增大(图6,7);礁间出现含砂鲕粒颗粒滩(图3f,h)也说明生物礁发育于水体能量不断增强的动荡环境。
-
一般来说,后生动物参与建造的生物礁形成于寡营养、温暖、清洁、透光好、相对稳定的浅海动荡环境[1]。以大河坝剖面为代表的寒武纪早期生物礁形成环境可能并非如此。虽然SiO2和CaO含量变化显示古杯动物与钙质微生物联合建礁体中陆源输入量处于仙女洞组沉积期最低水平(图7、图8b),但是详细的岩相学证据显示不仅层状薄礁体内含粉砂—黏土级陆源组分,不同期次礁体间也主要为陆源碎屑薄层。这些证据显示出古杯—钙质微生物联合建造具有一定的浑浊水环境耐受度(图4,5),可能与滤食性古杯动物对周围环境泥沙具有相对较强的适应性和忍耐力有关[15,25]。另一方面,细粒陆源碎屑组分输入带来的营养物质(如磷、铁等)[37]可提高生物礁生长的营养水平,促进初级生产力繁盛[47],也有利于古杯生物矿化作用和微生物有机矿化作用形成格架或结壳[48](图4)。但是,生物礁顶部及盖层中大量存在中砂级别的岩屑和石英颗粒,且SiO2含量快速增加(图7),显示陆源输入通量快速提高,最终造成造礁生物死亡后被持续的砂体堆积掩埋难以恢复。
-
大河坝剖面包括7种主要岩相类型,对应的沉积环境包括陆棚、过渡带、下临滨、上临滨、洪积扇、生物礁和鲕粒滩。仙女洞组沉积期整体处于从滨外向滨岸带演化的海退序列,生物礁发育于临滨带下部至上部过渡环境。生物礁发育经历了以凝块石主导,古杯动物主导,以及古杯与凝块石共同建造三个阶段。凝块石建造为生物礁的发育提供了硬质基底,礁体中上部由多套叠置的、具隆起外形的薄层古杯—钙质微生物联合堆积体构成,陆源碎屑组分常见。礁体生长与整体陆源输入的显著降低以及古杯动物与微生物对弱混浊环境的适应能力有关,即较低的陆源输入有利于古杯—钙质微生物礁的发育。但大量粗粒陆源组分的持续输入最终造成大河坝礁体被掩埋。
Petrographic Features and Paleoenvironmental Significance of the Lower Cambrian Reef in the Xiannüdong Formation, Mian County, Southern Shaanxi
-
摘要: 目的 生物礁演化史上一个关键节点是寒武纪早期广泛出现了后生动物古杯与微生物群落的联合造礁,该造礁群落形成环境和发育条件值得探讨。 方法 以华南板块西北缘汉南—米仓山地区的勉县大河坝剖面为例,对寒武系仙女洞组沉积序列和生物礁临近层位开展了系统的岩相学、沉积学和元素地球化学分析。 结果 岩相学特征显示礁体形成于海退背景下的临滨带环境,建造过程可依次分为钙质微生物主导、古杯动物大量繁盛以及古杯与钙质微生物联合建造三个阶段。生物礁灰岩中的陆源碎屑组分含量向上增多,且颗粒变粗,顶部被中—细砂岩覆盖。 结论 古杯—钙质微生物礁生长过程中对低强度的陆源碎屑输入具有一定的耐受度,当粗粒陆源碎屑组分的持续、高强度输入时将造成生物礁发育终止。本实例揭示了古杯—钙质微生物造礁群落生长过程与海底生态指标的相关性,可为古环境学、沉积学和储层地质学相关研究提供参考。
-
关键词:
- 古杯—钙质微生物群落 /
- 礁灰岩 /
- 古环境指标 /
- 寒武纪第三期 /
- 汉南—米仓山地区
Abstract: Objective A key point in the evolutionary history of reefs is the widespread occurrence of associated archaeocyath-calcimicrobial communities in the Early Cambrian. The developmental environments and conditions for these reef-building communities deserve more attention. Methods We present a case study of the Daheba section in Mian county, located in the Hanzhong-Micangshan area along the northwestern margin of the South China Block. In this region, we conducted comprehensive petrographic, sedimentological, and geochemical analysis of sedimentary successions to investigate reef development in turbid environments. Results A detailed petrographic study indicates that the Daheba reef was formed in a near-shore, high-energy condition. Calcified microfossils, archaeocyaths and archaeocyath-calcimicrobial communities dominated reef construction from the early to late phases of reef growth. Small amounts of terrigenous particles were found within the inner reef framework, increasing from the base to the top. Conclusions This indicates that the reef was tolerant of a small input of terrigenous silts and fine sands during growth. These terrigenous materials material may have provided a source of nutrients and facilitated reef growth; however, an excess of siliciclastics would have abraded reef-building organisms and blocked their feeding passages, perhaps even buried the reef architecture completely. This study has clarified the growth process and environment of an Early Cambrian archaeocyathan-calcimicrobial reef in China, and may provide a reference for further study of Early Cambrian reef systems in paleoenvironmental reconstruction, sedimentology and petroleum geology. -
图 1 研究区位置及寒武纪第三期古地理图
Figure 1. Location and paleogeography (Cambrian Age 3 ) of the study area

图 3 大河坝剖面仙女洞组露头
Figure 3. Photographs showing the outcrops characteristics of Xiannüdong Formation at the Daheba section
图 4 大河坝仙女洞组造礁古杯动物和钙质微生物抛光面特征
Figure 4. Polished slabs showing archaeocyaths and calcified microbes identified in the Xiannüdong Formation of the Daheba section
图 5 大河坝生物礁层的岩相学特征
Figure 5. Lithofacies of photomicrographs showing the petrological characteristics of the Daheba reef
图 6 生物礁内部陆源碎屑颗粒粒级及分布
Figure 6. Size distribution of terrigenous particles in the Daheba reef
图 7 大河坝剖面仙女洞组综合柱状图
Figure 7. Integrated characteristics of lithofacies, sea level, and major element contents at the Daheba section
表 1 大河坝剖面主要岩相类型和特征
Table 1. Lithofacies types and characteristics of the Daheba section
编号 主要特征 沉积构造 沉积环境 LF1 薄层粉砂质泥岩为主,少量为薄层泥岩夹粉砂岩。可见生物活动钻孔,几乎不含生物化石,有机质含量较高,可见沥青充填孔隙 水平层理、见生物钻孔 深水陆棚 LF2 薄层含钙粉砂岩为主,部分为含钙粉砂岩与泥岩/粉砂质泥岩互层,偶夹钙质细砂岩纹层。生物钻孔孔径一般为2 mm,常被钙质粗粉砂—细砂充填 水平层理、生物钻孔非常发育 过渡带 LF3 中薄层钙质岩屑细砂岩与钙质粉砂岩互层,部分为细砂岩夹粉砂岩。钙质组分含量较高,最高可达30%,多以胶结物的形式产出。见海绿石,呈次圆状,磨圆良好,偶见异地搬运鲕粒以及三叶虫化石碎片。陆源碎屑颗粒分选较好,磨圆中等—差;岩屑含量多于石英,次棱角状—次圆状 平行层理 下临滨 LF4 底部和顶部为中厚层含钙石英岩屑中砂岩,分选中等,颗粒之间呈点—线接触;钙质组分以胶结物形式充填陆源颗粒之间孔隙,含量一般不超过10%,且不含生物化石。中部为块状含砾岩屑粗砂岩,分选、磨圆极差,成分成熟度与结构成熟度低,并显示出杂乱堆积的特征。砾石粒径在2~5 mm,砾石间充填细砂至粗砂颗粒以及少量的泥,不含生物碎屑 递变层理、砾石叠瓦构造 洪积扇 LF5 块状含砂(砂质)生物灰岩和凝块石。其中含砂生物灰岩中陆源碎屑组分约为10%~20%,以岩屑为主,石英次之,颗粒分选中等,磨圆较差,多分布于古杯和钙质微生物周缘。古杯常与钙质微生物共生。钙质微生物类型为附枝菌和肾形菌,生物格架之间多为亮晶方解石胶结物。砂质凝块石中陆源碎屑组分含量超过30%,颗粒粒径可达粗粉砂级,分选较好,磨圆差,石英和岩屑颗粒多为次棱角状。古杯含量少。钙质微生物特征受高通量陆源物质混入影响,保存差 未见 生物礁 LF6 厚层含砂鲕粒灰岩。鲕粒粒径一般在0.6~1 mm,同心—放射状结构为主,部分被白云石晶粒交代;亮晶方解石胶结,分选好,可见少量古杯和三叶虫化石碎片。陆源碎屑组分主要为中砂和细砂级石英和岩屑颗粒,分选较好,磨圆中等—较差,颗粒含量占总体约10%~25% 未见 鲕粒滩 LF7 中层状含钙细砂岩和中—细砂岩。陆源碎屑组分分选良好,磨圆中等,颗粒类型包括岩屑(含量约40%)、石英(约50%)、长石(<10%)。颗粒之间呈凹凸接触,多为棱角—次棱角状。钙质组分含量不超过15%,主要为陆源碎屑颗粒胶结物形式,部分云化,未见碳酸盐颗粒 平行层理 上临滨  下载: 导出CSV
下载: 导出CSV
-
[1] James N P, Wood R. Reefs[M]//James N P, Dalrymple R W. Facies models 4. St. John's: Geological Association of Canada, 2010: 421-447. [2] Wood R. Reef evolution[M]. Oxford: Oxford University Press, 1999:1-426. [3] Armstrong A K, Snavely P D, Addicott W O. Porosity evolution of Upper Miocene reefs, Almeria Province, southern Spain[J]. AAPG Bulletin, 1980, 64(2): 188-208. [4] Sarg J F. Oil and gas reservoirs and coral reefs[M]//Hopley D. Encyclopedia of modern coral reefs. Encyclopedia of earth sciences series. Dordrecht: Springer, 2011: 745-751. [5] Webb G E. Was Phanerozoic reef history controlled by the distribution of non-enzymatically secreted reef carbonates (microbial carbonate and biologically induced cement)?[J]. Sedimentology, 1996, 43(6): 947-971. [6] Pratt B R. Early cementation and lithification in intertidal cryptalgal structures, Boca Jewfish, Bonaire, Netherlands Antilles[J]. Journal of Sedimentary Research, 1979, 49(2): 379-386. [7] Kirkland B L, Dickson J A D, Wood R A, et al. Microbialite and microstratigraphy: The origin of encrustations in the middle and upper Capitan Formation, Guadalupe Mountains, Texas and New Mexico, USA[J]. Journal of Sedimentary Research, 1998, 68(5): 956-969. [8] Wang X, Foster W J, Yan J X, et al. Delayed recovery of metazoan reefs on the Laibin-Heshan platform margin following the Middle Permian (Capitanian) mass extinction[J]. Global and Planetary Change, 2019, 180: 1-15. [9] Grotzinger J P, James N P. Precambrian carbonates: Evolution of understanding[M]//Grotzinger J P, James N P. Carbonate sedimentation and diagenesis in the evolving Precambrian world. Tulsa: SEPM, 2000: 3-20. [10] Li F, Deng J T, Kershaw S, et al. Microbialite development through the Ediacaran-Cambrian transition in China: Distribution, characteristics, and paleoceanographic implications[J]. Global and Planetary Change, 2021, 205: 103586. [11] Wood R A, Grotzinger J P, Dickson J A D. Proterozoic modular biomineralized metazoan from the Nama Group, Namibia[J]. Science, 2002, 296(5577): 2383-2386. [12] Penny A M, Wood R A, Curtis A, et al. Ediacaran metazoan reefs from the Nama Group, Namibia[J]. Science, 2014, 344(6191): 1504-1506. [13] Mehra A, Maloof A. Multiscale approach reveals that Cloudina aggregates are detritus and not in situ reef constructions[J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(11): E2519-E2527. [14] Yang B, Steiner M, Schiffbauer J D, et al. Ultrastructure of Ediacaran cloudinids suggests diverse taphonomic histories and affinities with non-biomineralized annelids[J]. Scientific Reports, 2020, 10(1): 535. [15] Zhuravlev A Y. Paleoecology of Cambrian reef ecosystems[M]//Stanley G D. The history and sedimentology of ancient reef systems. Boston: Springer, 2001: 121-157. [16] Hicks M, Rowland S M. Early Cambrian microbial reefs, archaeocyathan inter-reef communities, and associated facies of the Yangtze Platform[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2009, 281(1-2): 137-153. [17] Adachi N, Nakai T, Ezaki Y, et al. Late Early Cambrian archaeocyath reefs in Hubei province, South China: Modes of construction during their period of demise[J]. Facies, 2014, 60(2): 703-717. [18] Cordie D R, Dornbos S Q, Marenco P J. Evidence for a local reef eclipse in a shallow marine carbonate environment following the regional extinction of archaeocyaths in Laurentia (stage 4, Cambrian)[J]. Facies, 2020, 66: 5. [19] Pruss S B, Smith E F, Leadbetter O, et al. Palaeoecology of the archaeocyathan reefs from the Lower Cambrian Harkless Formation, southern Nevada, western United States and carbon isotopic evidence for their demise[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2019, 536: 109389. [20] Cordie D R, Dornbos S Q, Marenco P J, et al. Depauperate skeletonized reef-dwelling fauna of the Early Cambrian: Insights from archaeocyathan reef ecosystems of western Mongolia[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2019, 514: 206-221. [21] Álvaro J J, Clausen S, El Albani A, et al. Facies distribution of the Lower Cambrian cryptic microbial and epibenthic archaeocyathan-microbial communities, western Anti-Atlas, Morocco[J]. Sedimentology, 2006, 53(1): 35-53. [22] Lee J H, Chen J T, Chough S K. The Middle-Late Cambrian reef transition and related geological events: A review and new view[J]. Earth-Science Reviews, 2015, 145: 66-84. [23] Kiessling W. Secular variations in the Phanerozoic reef ecosystem[M]//Kiessling W, Flügel E, Golonka J. Phanerozoic reef patterns. Tulsa: SEPM, 2002: 625-690. [24] 张俊明,袁克兴. 湖北宜昌王家坪下寒武统天河板组古杯礁丘及其成岩作用[J]. 地质科学,1994,29(3):236-245. Zhang Junming, Yuan Kexing. Archaeocyath reefs from the Lower Cambrian Tianheban Formation at Wangjiaping, Yichang, Hubei and their diagenesis[J]. Scientia Geologica Sinica, 1994, 29(3): 236-245. [25] Wood R A, Zhuravlev A Y, Debrenne F. Functional biology and ecology of archaeocyatha[J]. Palaios, 1992, 7(2): 131-156. [26] Zhuravlev A Y. Reef ecosytem recovery after the Early Cambrian extinction[J]. Geological Society, London, Special Publications, 1996, 102(1): 79-96. [27] 袁克兴,朱茂炎,张俊明,等. 陕西南郑福成剖面早寒武世古杯生物地层学:对古杯演化和地层对比的初步探讨[J]. 古生物学报,2001,40(增刊1):115-129. Yuan Kexing, Zhu Maoyan, Zhang Junming, et al. Biostratigraphy of archaeocyathan horizons in the Lower Cambrian Fucheng section, south Shaanxi province: Implications for regional correlations and archaeocyathan evolution[J]. Acta Palaeontologica Sinica, 2001, 40(Suppl.1): 115-129. [28] 袁克兴,章森桂. 华中—西南区早寒武世古杯化石[J]. 古生物学报,1980,19(5):380-392,433-436. Yuan Kexing, Zhang Sengui. Lower Cambrian archaeocyatha of central and southwestern China[J]. Acta Palaeontologica Sinica, 1980, 19(5): 380-392, 433-436. [29] 杨爱华. 扬子地台早寒武世古杯动物群及其灭绝事件[D]. 南京:中国科学院研究生院,2005:1-234. Yang Aihua. The Early Cambrian archaeocyaths and their extinction event on the Yangtze Platform[D]. Nanjing: Graduate School of Chinese Academy of Sciences, 2005:1-234. [30] 刘仿韩,苏春乾,杨友运,等. 米仓山南坡寒武系沉积相分析[J]. 西安地质学院学报,1987,9(4):1-12. Liu Fanghan, Su Chunqian, Yang Youyun, et al. Sedimentary facies analysis of Cambrian in the south of Micang Mountain[J]. Journal of Xi’an College of Geology, 1987, 9(4): 1-12. [31] 张廷山,兰光志,沈昭国,等. 大巴山、米仓山南缘早寒武世礁滩发育特征[J]. 天然气地球科学,2005,16(6):710-714. Zhang Tingshan, Lan Guangzhi, Shen Zhaoguo, et al. Early Cambrian reefs and banks development in southern margin of Daba Mt. and Micang Mt.[J]. Natural Gas Geoscience, 2005, 16(6): 710-714. [32] Yang A H, Zhu M Y, Zhuravlev A Y, et al. Archaeocyathan zonation of the Yangtze Platform: Implications for regional and global correlation of Lower Cambrian stages[J]. Geological Magazine, 2016, 153(3): 388-409. [33] 杨慧宁,毛颖颜,潘兵,等. 陕南寒武纪早期仙女洞组生物礁灰岩微相序列[J]. 微体古生物学报,2016,33(1):75-86. Yang Huining, Mao Yingyan, Pan Bing, et al. Microfacies sequences of the early Cambrian (Series 2) Xiannüdong Formation reefs in southern Shaanxi province, NW China[J]. Acta Micropalaeontologica Sinica, 2016, 33(1): 75-86. [34] Zhang M Q, Hong J, Choh S J, et al. Thrombolite reefs with archaeocyaths from the Xiannüdong Formation (Cambrian Series 2), Sichuan, China: Implications for Early Paleozoic bioconstruction[J]. Geosciences Journal, 2017, 21(5): 655-666. [35] Tang H, Kershaw S, Tan X C, et al. Sedimentology of reefal buildups of the Xiannüdong Formation (Cambrian series 2), SW China[J]. Journal of Palaeogeography, 2019, 8: 11. [36] 曾楷,李飞,龚峤林,等. 寒武系第二统仙女洞组混合沉积特征及古环境意义:以川北旺苍唐家河剖面为例[J]. 沉积学报,2020,38(1):166-181. Zeng Kai, Li Fei, Gong Qiaolin, et al. Characteristics and paleoenvironmental significance of mixed siliciclastic-carbonate sedimentation in the Xiannüdong Formation, Cambrian (Series 2): A case study from the Tangjiahe section, Wangcang, northern Sichuan[J]. Acta Sedimentologica Sinica, 2020, 38(1): 166-181. [37] Li H, Li F, Li X, et al. Development and collapse of the Early Cambrian shallow-water carbonate factories in the Hannan-Micangshan area, South China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2021, 583: 110665. [38] 邓嘉婷,李飞,龚峤林,等. 埃迪卡拉纪—寒武纪之交微生物岩特征对比及古海洋学意义:以汉南—米仓山地区为例[J]. 古地理学报,2021,23(5):919-936. Deng Jiating, Li Fei, Gong Qiaolin, et al. Characteristics and palaeoceanographic significances of microbialite development in the Ediacaran-Cambrian transition: A case study from Hannan-Micangshan area[J]. Journal of Palaeogeography (Chinese Edition), 2021, 23(5): 919-936. [39] 李红,李飞,龚峤林,等. 混积岩中重矿物形貌学特征及物源意义:以川北寒武系第二统仙女洞组为例[J]. 沉积学报,2021,39(3):525-539. Li Hong, Li Fei, Gong Qiaolin, et al. Morphological characteristics and provenance significance of heavy minerals in the mixed siliciclastic-carbonate sedimentation: A case study from the Xiannüdong Formation, Cambrian (series 2), northern Sichuan[J]. Acta Sedimentologica Sinica, 2021, 39(3): 525-539. [40] Dong Y P, Liu X M, Santosh M, et al. Neoproterozoic accretionary tectonics along the northwestern margin of the Yangtze Block, China: Constraints from zircon U-Pb geochronology and geochemistry[J]. Precambrian Research, 2012, 196-197: 247-274. [41] Peng S C, Babcock L E, Ahlberg P. The Cambrian period[M]//Gradstein F M, Ogg J G, Schmitz M D, et al. Geologic time scale 2020. Amsterdam: Elsevier, 2020: 565-629. [42] 朱茂炎,孙智新,杨爱华,等. 中国寒武纪岩石地层划分和对比[J]. 地层学杂志,2021,45(3):223-249. Zhu Maoyan, Sun Zhixin, Yang Aihua, et al. Lithostratigraphic subdivision and correlation of the Cambrian in China[J]. Journal of Stratigraphy, 2021, 45(3): 223-249. [43] 张文堂,袁克兴,周志毅,等. 西南地区的寒武系[M]//中国科学院南京地质古生物研究所. 西南地区碳酸盐生物地层. 北京:科学出版社,1979:39-107. Zhang Wentang, Yuan Kexing, Zhou Zhiyi, et al. Cambrian of southwest China[M]//Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences. Biostratigraphy of carbonates of Southwest China. Beijing: Science Press, 1979: 39-107. [44] Shapiro R S. A comment on the systematic confusion of thrombolites[J]. Palaios, 2000, 15(2): 166-169. [45] Dupraz C, Strasser A. Nutritional modes in coral-microbialite reefs (Jurassic, Oxfordian, Switzerland): Evolution of trophic structure as a response to environmental change[J]. Palaios, 2002, 17(5): 449-471. [46] 吴亚生,姜红霞,李莹,等. 微生物碳酸盐岩的显微结构基本特征[J]. 古地理学报,2021,23(2):321-334. Wu Yasheng, Jiang Hongxia, Li Ying, et al. Microfabric characteristics of microbial carbonates[J]. Journal of Palaeogeography (Chinese Edition), 2021, 23(2): 321-334. [47] Gerdes G. What are microbial mats?[M]//Seckbach J, Oren A. Microbial mats: Modern and ancient microorganisms in stratified systems. Dordrecht: Springer, 2010: 3-25. [48] 李杨凡,李飞,王夏,等. 上扬子北缘寒武纪早期后生动物礁特征及古环境意义[J]. 地球科学,2023,48(11):4321-4334. Li Yangfan, Li Fei, Wang Xia, et al. Sedimentary Characteristics and Paleoenvironmental Significance of Early Cambrian Metazoan Reefs in Northern Margin of Upper Yangtze Block[J]. Earth Science, 2023, 48(11): 4321-4334. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 765
- HTML全文浏览量: 267
- PDF下载量: 64
- 被引次数: 0

