

 下载:
下载:



-
“碳酸盐(生产)工厂”(carbonate factory)是近年来碳酸盐岩沉积学领域研究的一个重要方向,其强调了生物作为主要建造者在自身和外部环境因素影响下如何调控规模性碳酸盐沉积物(岩)的形成过程,在研究生物与环境因素对沉积体系的影响、台地建造样式和演化过程,以及碳酸盐岩有利储集相带预测等方面具有重要价值[1⁃4]。“碳酸盐(生产)工厂”早期被定义为大规模碳酸盐沉积物汇聚的场所,既包括沉积物的主要产地——潮下带,也包括分别向岸和向盆地方向搬运后保存有碳酸盐沉积记录的滨岸和斜坡环境[5]。“碳酸盐(生产)工厂”在大规模碳酸盐形成与保存的过程中会受到一系列因素的调控,如水温、光照条件、浊度、营养水平、水动力条件、盐度、海平面变化、碳酸盐矿物饱和度和海水化学条件等[6⁃12]。Schlager[12⁃14]在前人一系列工作的基础上总结、归纳出三种主要碳酸盐生产工厂类型,即:(1)热带浅水工厂(tropical shallow-water factory),主要由光合自养生物及其共生生物构成,形成于光照条件良好的热带暖水环境;(2)温凉水工厂(cool-water factory),主要为异养生物群落构成,多集中于光照条件差,营养丰富的温、冷水环境;(3)灰泥丘工厂(mud-mound factory),主要为生物诱导矿化和无机沉淀方式结合的原地细粒碳酸盐构成,一般形成于富营养环境。之后,Schlager[12]对工厂体系又进一步完善,用“T工厂”指代“热带(tropical)”和“水体上部(top-of-the-water-column)”工厂类型,用“C工厂”代表“温凉水(cool-water)”和“受控型沉淀过程(controlled-precipitation)”工厂类型,以及用“M工厂”代表“灰泥丘(mud-mound)”“泥晶(micrite)”和“微生物(microbes)”参与的工厂类型。其中,“M工厂”在地球上出现时间最早[15],主要通过有机矿化过程调控碳酸盐生产[16],是本文主要关注工厂类型。考虑到“M工厂”包括三种不同的碳酸盐生产方式,而本文主要强调底栖微生物群落在“M工厂”形成过程中的作用,因而在后续论述中将其细化为微生物碳酸盐生产工厂。
微生物碳酸盐生产工厂指由底栖微生物群落作为主要建造者,通过黏结、捕获作用和有机矿化作用形成碳酸盐沉积物的环境、作用方式和建造样式的总称。构成微生物碳酸盐生产工厂的样式按照Grey et al.[17]最新方案包括叠层石、凝块石、树枝石、均一石和微生物诱导沉积构造(Microbial Induced Sedimentary Structures,MISS);共生的其他碳酸盐沉积物包括球粒、内碎屑、鲕粒和真核生物等[18]。微生物碳酸盐生产工厂发育非常广泛,在海洋及湖泊浅水至深水环境均可发育[19⁃20],本文着重介绍其在海洋环境下的特征。受水深条件和营养水平的影响,现代海洋环境下微生物碳酸盐生产工厂主要在浪基面之下的中等水深条件发育[14],在一些正常或相对局限的浅水环境中也存在一些实例[21⁃22]。地质历史时期,微生物碳酸盐生产工厂在前寒武纪占主导地位[23⁃24],随着后生动物的勃发在浅水环境出现明显衰退[25⁃28],但在一些极端事件(如生物绝灭事件后)或局限环境仍可在浅水环境广泛发育[29⁃30]。现代最为典型的微生物碳酸盐生产工厂保存在澳大利亚鲨鱼湾Hamelin Pool[18];该地区是一个局限海湾,拥有极高的碳酸盐矿物饱和度和盐度,从潮下至潮上环境发育类型、丰度、特征、内部结构各异的微生物碳酸盐及其共生建造[22],是一个研究程度极高的现代实例[18,22,31⁃33]。虽然微生物碳酸盐生产工厂在地质历史时期的分布非常广泛,但是研究程度普遍较低,已有研究主要集中在微生物岩的时空演化序列和对极端事件响应方面[34⁃36],对工厂自身发育过程和影响因素方面的研究较为匮乏。考虑到地质历史时期存在大量微生物碳酸盐沉积且与环境演化关系密切,厘清微生物碳酸盐生产工厂的建造过程和调控因素是一个值得探索的重要科学问题。
本文以河南云梦山剖面寒武系崮山组为例,对苗岭世古丈期一个典型的Hamelin Pool型微生物碳酸盐生产工厂进行了解译,通过详细刻画不同工厂样式的组成和发育特征,厘清其形成过程和发育环境,探索微生物碳酸盐生产工厂的启动和关闭机制,为深时微生物碳酸盐生产工厂研究提供一个可对比的研究实例。相关工作的开展有利于了解不同类型碳酸盐生产工厂的建造样式和影响因素,并在理解寒武纪海洋浅水生态系统和沉积过程、微生物群落环境适应性以及有机矿化机制等方面具有一定的参考价值。
-
华北地区在寒武纪为一个位于低纬度地区的独立地块,在新元古代至寒武纪早期“大不整合”事件之后开始接受沉积[37]。受寒武纪第二世晚期大规模海侵事件的影响,华北板块从周缘向内部逐渐被海水覆盖,形成广阔的陆表海;板块整体表现为西高东低、南高北低的古地理格局,且以潮坪和潟湖环境为主[38]。苗岭世时,除鄂尔多斯古陆和鲁西古陆外,其他地区大部分被海水覆盖,从陆地向深水方向依次发育了潟湖、潮坪、浅滩、缓坡和陆棚环境,以清水碳酸盐沉积为主(图1)。至芙蓉世时,华北板块受海退影响,碳酸盐沉积物不断向较深水区进积;浅水环境主要为开阔台地,古陆周缘则以局限台地为主[38]。

图 1 华北寒武纪苗岭世古地理图及研究区岩石地层单元划分
Figure 1. Palaeogeography of the North China Block in the Cambrian Miaolingian, and the lithostratigraphic column of the study area
华北寒武系出露良好,从下至上依次为辛集组、朱砂洞组、馒头组、张夏组、崮山组、炒米店组,及部分三山子组(穿时),一些地区寒武系底部缺失辛集组[39]。研究区位于河南省鹤壁市淇县与新乡市卫辉交界的云梦山景区内,寒武系朱砂洞组至炒米店组发育完整,出露情况良好(图1)。本次研究关注的崮山组属苗岭统古丈阶,厚度近30 m,底部以三叶虫Blackwelderia开始出现为标志[39⁃40],岩性主要为灰色薄—中层条带状灰岩、砾屑(竹叶状)灰岩、泥晶灰(白云)岩、鲕粒灰岩,以及中—厚层微生物岩。崮山组下伏地层为张夏组,岩性以灰色中—厚层鲕粒灰岩为主,含少量微生物岩和砾屑灰岩;上覆地层为炒米店组,岩性主要为中—厚层细晶白云岩、瘤状白云岩,以及条带灰岩、含生物遗迹灰岩和生物扰动灰(白云)岩[39⁃41]。
-
目前对微生物碳酸盐生产工厂的研究较为薄弱,还没有广为接受的研究方法和实施方案供参考。研究尝试从工厂生产定量化评价的角度出发,主要通过系统的野外观察和统计学工作,结合光面特征对典型微生物岩类型、内部结构和生长过程进行精细刻画,对工厂发育的环境因素和自身建造过程进行分析。对微生物岩形貌特征观察采用广泛接受的Shapiro[42]方案,主要对它的宏观结构、大型结构和中观结构进行区分。对微生物岩结构特征描述采用Grey et al.[17]方案(图2)。野外采用网格法定量统计相关参数,步骤包括:(1)对每个微生物岩露头垂向上按横纵坐标每10 cm间隔标记并拍摄大量细节照片,完成详细观察描述;(2)室内将照片整理后利用CorelDRAW软件绘制长、宽均为1 cm的密集网格线;(3)根据野外描述记录统计网格线内不同微生物组构宽度、高度、类型占比以及其他参与工厂建造的颗粒类型和分布情况等参数,辅以少量光面和薄片进行微观特征验证。
-
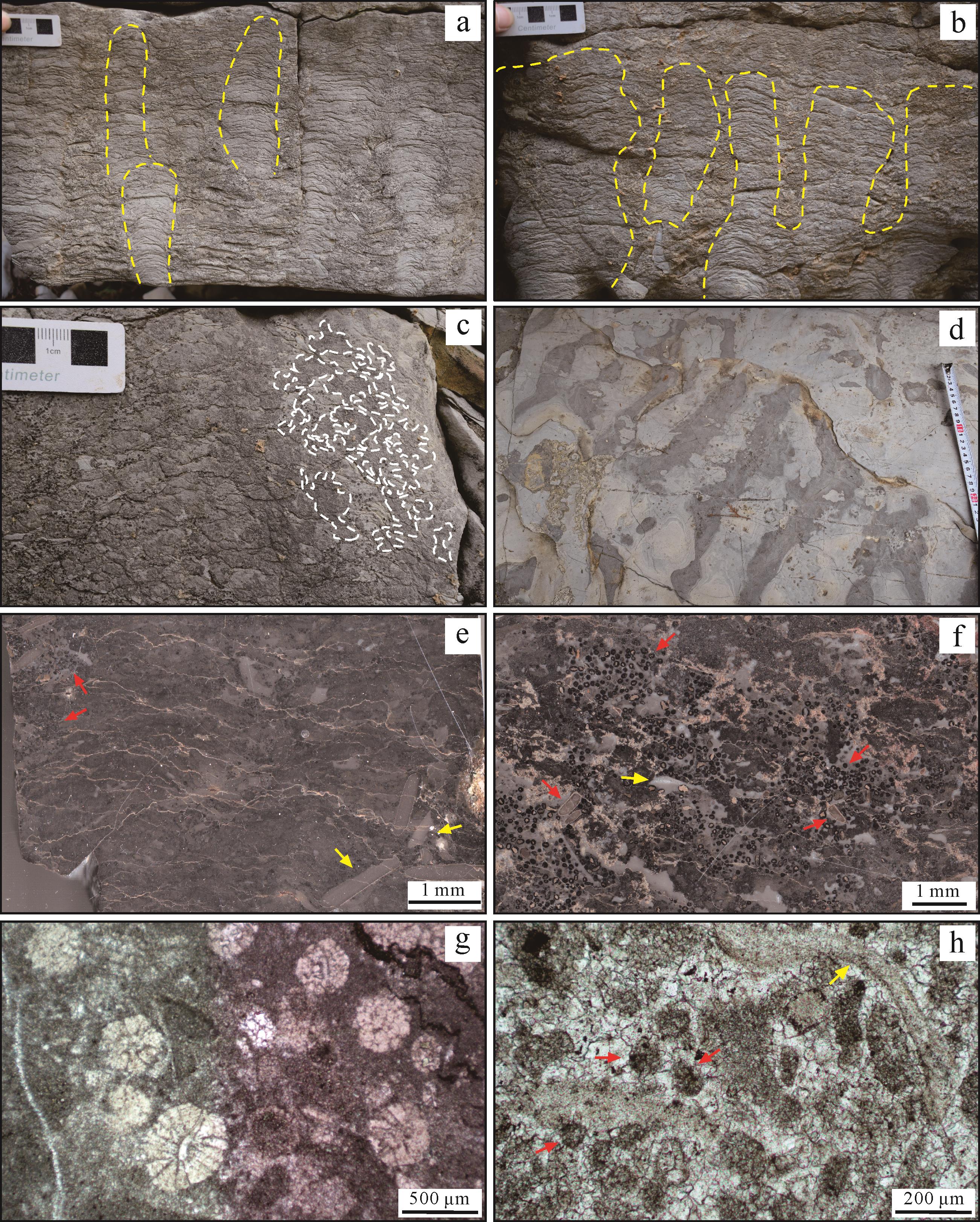
研究区崮山组中记录的微生物碳酸盐生产工厂的构成具有特殊性。工厂构成的基本单元包括叠层石、凝块石、树枝石、均一石和MISS构造(图3)。这些单元既可以单独产出(图3a~f),也可在空间上相互叠置形成共生关系(图4),保存的整体形态有近球状(图4a~c)、大型板状(图4d~f)和穹窿状(图4g~i)等类型。叠层石产出类型多样,单体形态包括柱状、穹窿状和层状等,这些单体常构成复合结构(图4)。层状结构叠层石内部一般具波状起伏的纹层(图4a,b);穹窿状叠层石少见,主要为近球状(图4a);柱状叠层石常见,以条带状泥晶灰岩为基底(图4a,d),部分以层状和穹窿状叠层石为基底,柱体以直立状、分枝状、汇聚状和交织状等较为常见(图3a,b、图4a,b,d,e)。凝块石主要为小型穹窿状或层状形式产出(图3c、图4a),以次圆形、朵形凝块为主(图4e,h)。树枝石主要发育于大型穹窿状结构的内部,凝块呈辐射的树枝状结构(图4g,h)。均一石形成于大型穹窿状结构的顶部,呈薄层状,内部成分均匀,与下伏微生物组构紧密接触而与上覆岩层存在截然界面(图4a,b),这与泥晶灰岩自然层存在明显差别(图4d,e)。MISS构造主要发育于大型穹窿状微生物岩层顶面,保存在波痕的波脊处,波峰和波谷均较为圆滑,波痕高度不超过3 cm,具典型流水波痕特点(图3d)。其他参与者还包括鲕粒、内碎屑、灰泥,以及少量生物碎屑(图3e~h)。鲕粒主要保存在微生物层状结构和凝块结构内(图3e),或者结构之间的孔洞(图3f,g),显示出异地埋藏特征。球粒粒径主要介于50~80 μm,磨圆极好,内部为泥晶化颗粒(图3h)。内碎屑颗粒丰富,主要保存在工厂基本单元周围,分选较差,磨圆较好(图3e、图4a)。工厂发育的基底主要为被侵蚀的条带状泥晶灰岩,向上突出具隆起正地形(图4);盖层主要为具典型超覆特征的鲕粒泥粒—颗粒灰岩(图4a),或者薄层泥晶灰岩(图4d,g)。

图 3 研究区崮山组微生物碳酸盐生产工厂岩石学特征
Figure 3. Photographs showing the lithological characteristics of the microbial carbonate factory preserved in the Gushan Formation

图 4 研究区典型微生物碳酸盐生产工厂复合结构类型和主要特征
Figure 4. Field photographs of the composite structures preserved within the microbial carbonate factory, with corresponding sketch maps and grid⁃based statistics
-
微生物碳酸盐生产工厂在崮山组成层性好,侧向延伸一般超过10 m,单层厚度一般不超过1 m,具旋回发育特点;间隔层位岩性主要为薄层鲕粒岩或泥晶灰岩。单层内工厂的基本单元高度大致相同,基底类型(条带灰岩为主)相似,但各个工厂单元样式和形貌具差异性(图4)。现将三种典型的工厂建造形式详细介绍如下。
-
特征描述:叠层石—凝块石—均一石复合结构发育于崮山组下部,整体呈近球状(图4a,b),高43 cm,宽51 cm,基底为条带状灰岩,向上依次为叠层石、凝块石和均一石,盖层为鲕粒灰岩。其中基底形状不规则,高约12 cm,宽约20 cm,被侵蚀改造特征明显。叠层石构成复合结构的主体,占比面积61%(以下占比面积均指占整个微生物岩的垂向组构面积),高33 cm,宽51 cm,由三个主要的柱体构成,彼此在工厂发育早期相互独立,晚期有一些纹层的汇聚(图4b)。柱体间常见鲕粒和内碎屑颗粒,以及一些发育规模较小的柱状叠层石或者由破碎叠层石构成的角砾,分选、磨圆极差;柱体间组构占比面积18%。凝块石呈穹窿状,高5 cm,宽51 cm,占比面积14%;表面凹凸不平,主要为深色凝块状结构残余物。单个凝块为次圆形、椭圆形和朵形,凝块间可见鲕粒。顶部为帽状均一石,占比面积7%,高8 cm,宽50 cm,内部主要为泥晶组构,生物钻孔丰富(图4a,红色箭头所示)。盖层为鲕粒灰岩,沿复合结构两侧具典型超覆特征(图4a,黄色箭头所示)。
环境解译:作为基底的泥晶条带灰岩中灰泥占主导,推测形成于潮下带低能环境,但条带灰岩被侵蚀、改造特征明显,周围为大量竹叶状砾屑,推测当时受到了风暴浪改造。叠层石以大型柱状结构为主,具抗浪格架,柱体间为丰富的竹叶状内碎屑,推测其形成于潮下带高能环境。上覆凝块石不具有格架结构,且凝块间主要为灰泥和异地搬运鲕粒,与叠层石组构相比,形成环境水体能量减弱,推测形成于潮间带。均一石内部主要为泥晶组构,形成水体能量最低,推测形成于潮间带上部至潮上带。随后,该样式受海侵过程中鲕粒超覆。该复合结构与现代澳大利亚鲨鱼湾Hamelin Pool潮坪环境微生物碳酸盐生产工厂特征非常相似 [22]。整体来看,该类型工厂形成于潮坪环境水体能量降低的变浅序列,随后被海侵初期的鲕粒滩所掩盖。
-
特征描述:叠层石—凝块石板状复合结构同样发育于崮山组下部,整体呈板状(图4d~f),高60 cm,可见宽度260 cm,存在一系列顺层发育的板状复合结构。该工厂基底为条带状灰岩和砾屑灰岩,主体为柱状叠层石,上部为穹窿状或板状凝块石。顶部虽然同样发育泥晶灰岩,但其在整个自然层顶部均有出现,并未与下伏板状复合结构构成整体,因此本文将这种类型的泥晶灰岩视为盖层而非均一石。其中,微生物碳酸盐生产工厂发育基底为平台状泥晶灰岩,宽60 cm,高15 cm,两端隆起呈脊状,内部可见大量砾屑堆积(高5 cm,宽6 cm),边缘破碎特征较为明显(图4e)。多个柱状叠层石以平台状泥晶灰岩为基底,形成宽60 cm,高40 cm的厚层,面积占比60%(图4f)。柱状叠层石在工厂发育早期较为孤立(高25~30 cm),晚期常发生汇聚形成大型交织状结构(图4d,e)。叠层石柱体之间孔隙主要被鲕粒、球粒和砾屑颗粒充填,面积占比14%。凝块石宽60 cm,高9 cm,面积占比26%,以深色椭圆形和朵形凝块为特征,凝块彼此相连构成支架,凝块间主要为分散的鲕粒和灰泥(图4d;因抗风化能力较弱而发生内凹)。
环境解译:该类型与叠层石—凝块石—均一石近球状复合结构具有一定相似性。其中基底条带状泥晶灰岩被波浪或风暴浪侵蚀、搅动特征明显,推测形成于易受风暴浪影响的潮下带环境。柱状叠层石的大量发育,以及柱体间充填的砾屑和鲕粒特征,推测形成于潮下带强水动力环境。上覆穹窿状凝块石层薄,推测形成于潮间带。泥晶灰岩作为盖层,可能指示了水体突然加深造成工厂被淹没。
-
特征描述:叠层石—树枝石—凝块石穹隆状复合结构发育于崮山组下部(图4g~i),高50 cm,宽180 cm。该复合结构纵向上以塔状外形的条带状泥晶灰岩为基底,宽60 cm,高15 cm。基底较陡侧主要发育树枝石,呈辐射状,宽30 cm,高20 cm,面积占比24%(图4g,h)。基底正上方发育层状叠层石,较缓侧基底主要为大量砾屑堆积,向上发育柱状叠层石并与基底正上方叠层石汇聚形成半球状;宽70 cm,高30 cm,面积占比28%。靠近基底的柱状叠层石紧密相连(宽度小于5 cm),远离基底的叠层石较为粗壮(近10 cm)。凝块石与树枝石接触关系较为复杂,从野外特征上看两者可能存在渐变和相互转化关系(图4g)。凝块石主要发育于半球状结构的上部和相对较缓一侧,单个凝块以近圆形、椭圆形和披针形为主,面积占比47%。凝块石被上覆泥晶灰岩覆盖。在穹隆状复合结构内部可见少量的鲕粒、球粒和砾屑组分,面积占比约1%。
环境解译:根据(1)基底泥晶灰岩被改造、侵蚀形成塔状结构,(2)基底内部为受软沉积变形和压实作用影响的破碎结构,以及(3)基底缓坡处为灰砾岩(砾屑灰岩)等特征,推测该工厂发育于受风暴强烈侵蚀、改造后的浅水环境。树枝石和叠层石的发育可能指示了潮下带上部强水动力条件。随后,板状凝块石发育于穹隆状复合体之上,指示了水体能量的进一步降低,推测其形成于潮间带环境。泥晶灰岩盖层指示了在海水快速加深过程中工厂发育停止。基底缓坡与陡坡环境沉积物类型和工厂样式存在明显不同,其原因可能与底形差异和水动力条件有关。
-
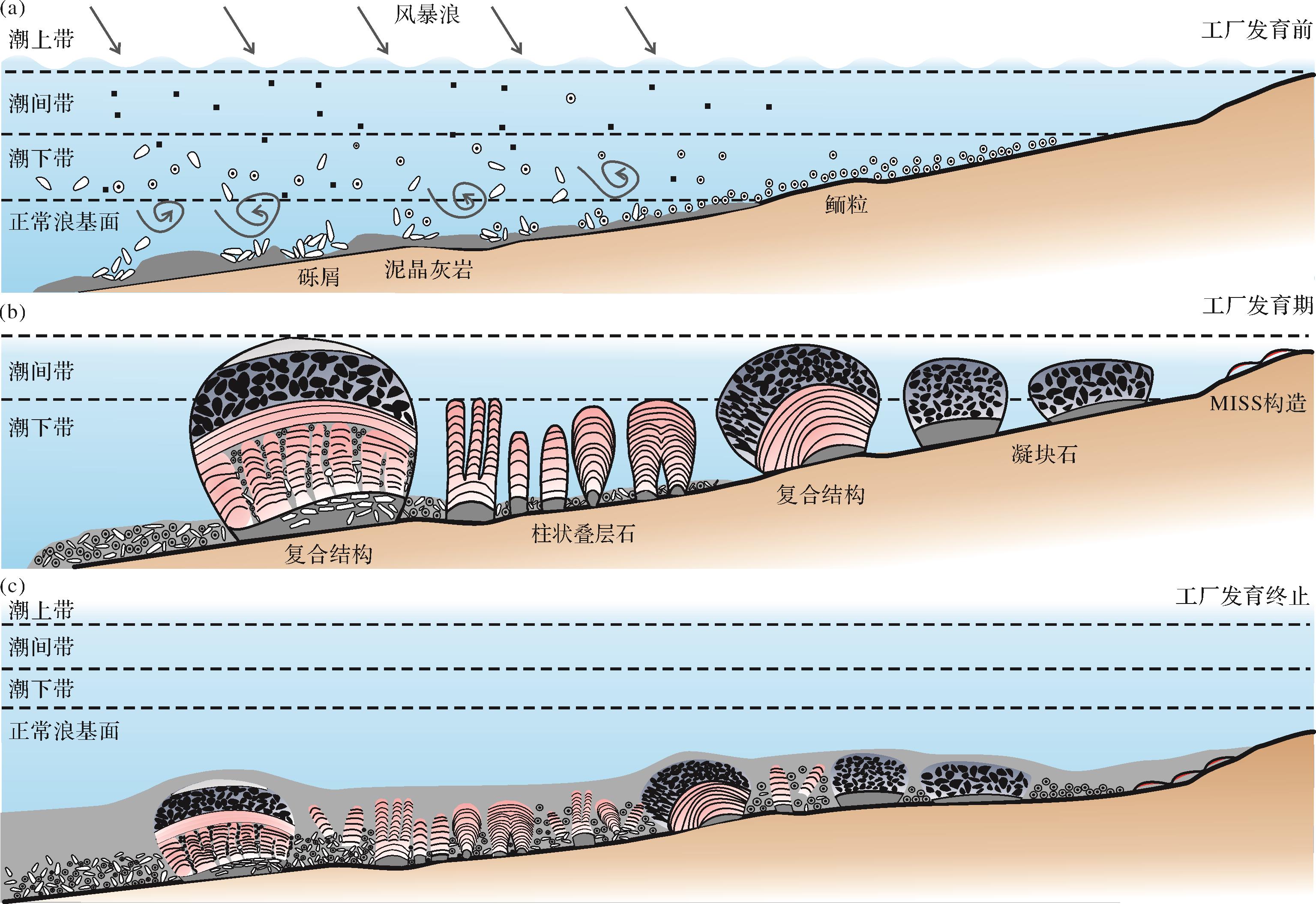
河南云梦山地区寒武纪苗岭世古丈期微生物碳酸盐生产工厂是深时工厂研究的一个典型实例。从构成上看,微生物碳酸盐生产工厂的建造样式包括叠层石、凝块石、树枝石、均一石和MISS构造,涵盖微生物岩的主要类型。同时,其建造样式既存在柱状叠层石等小型单独建造,还存在叠层石+凝块石+均一石、叠层石+树枝石+凝块石、叠层石+凝块石等复合类型(图4,5)。工厂发育基底主要为条带状泥晶灰岩,且泥晶灰岩往往形成突出底形,如平台状和塔状;盖层大部分为薄层泥晶灰岩,少量为鲕粒灰岩(图4)。微生物碳酸盐生产工厂主要保存在厚层灰岩层内(一般为50~100 cm),与泥晶灰岩或鲕粒灰岩间隔发育,从目前工作来看至少存在三个期次。另外,工厂发育范围较大,单期工厂横向延伸距离超百米,整体分布面积估计超1 km2。

图 5 研究区微生物碳酸盐生产工厂发育模式图
Figure 5. Development of the microbial carbonate factory (Guzhangian, Miaolingian) in the study area
从内部特征来看,研究区与现代Hamelin Pool微生物碳酸盐生产工厂具有较高的相似性。从构成上看,云梦山崮山组保存的微生物岩以柱状叠层石为主,类型包括圆柱状、圆筒状、鼻甲状(图3a,b),且单枝生长过程中可分叉、汇聚,甚至形成交织状结构(图4d,e)。凝块石层总体厚度很少超过30 cm,平铺板状外形较为常见,内部常保存一些异地埋藏的鲕粒和生屑(图3c,f)。树枝石发育较为少见,可见其形成大型辐射状结构,且与柱状叠层石相邻发育,显示出其较强的抗浪能力(图4g、图5b)。均一石往往呈现帽状外形和较为均匀的泥晶组构(图4a)。整体来看,这些特征与现代Hamelin Pool潮坪环境微生物碳酸盐生产工厂非常相似。与现代实例相比,云梦山工厂启动环境可能更为恶劣,风暴搅动特征明显(大量砾屑和风暴侵蚀改造)(图5a);大型半球状、穹窿状和板状建造启动于盛行风暴的间歇期(图5b)。依据柱状叠层石、凝块石和均一石(不普遍)沉积序列,抗浪能力逐渐降低的特点,推测微生物碳酸盐生产工厂形成于海退环境。
-
影响碳酸盐生产工厂发育的因素众多,包括温度、盐度、碱度、碳酸盐饱和度、光照条件、水动力条件、营养水平,以及海水化学性质(如Mg/Ca)等[43]。温度是调控生物生存和分布的主要因素之一,暖水环境有利于降低生物新陈代谢活动的能量损耗[12],也有利于生物钙化和维持非生物碳酸盐颗粒形成所需的高碳酸盐矿物饱和度条件[12,44]。同时,温度与碳酸盐矿物饱和度具有较好的线性关系[45],因而暖水条件下饱和度也可以维持在较高水平[9]。盐度条件指利用生物对盐度的适应性差异(广盐性和窄盐性)调控生物群落组合分布,特别是在高盐度环境下通过抑制窄盐性生物生存而为微生物群落繁盛创造较好的环境条件[12,46]。碱度通过调节水体中碳酸根和碳酸氢根等离子浓度影响生物骨骼钙化和方解石补偿深度[47],间接调控生物分布范围。另外,高碱度条件有利于微生物钙化结构的保存[48]。光照指生物主要通过光合作用调控碳酸盐产率,进而影响生态系统构成。因此透光性的差别会影响生物和群落类型[1,11],但以异养细菌为主导的底栖微生物群落与光照条件无大的关联[43]。水动力条件则通过影响颗粒磨圆和颠选、抗浪格架的构成和规模,以及灰泥的形成和搬运等形式影响工厂类型和构成[46]。另一方面,水动力条件会显著影响工厂发育的基底形态(图4,5),这也是制约工厂发育稳定性和规模的重要因素。特别是风暴浪,不仅对海底地形进行侵蚀改造,还通过搅动、搬运等方式影响水体的营养水平和含氧量[49⁃51]。营养水平是调控水体初级生产力的主要因素,一般与陆源输入增强和上升流等带来的初级生产力的提高有关[52]。较高的营养水平刺激浮游微生物的繁盛造成底栖生物生存环境光照受限;同时初级生产力繁盛后,有机质降解过程中耗氧量增加进而制约了水体溶解氧水平,因此可认为营养条件是制约工厂类型的关键因素之一[53⁃54]。最后,海水化学性质对碳酸盐生产工厂的影响主要体现在长周期尺度上,通过影响参与工厂建造的非骨屑颗粒、重度钙化(hypercalcifying)生物,以及胶结物矿物性质的方式调控生产者类型和建造速率[9,11,55]。
研究区微生物碳酸盐生产工厂的繁盛可能是当地高营养条件主导下,强水动力条件以及相对局限、咸化水体性质等方面综合作用的结果。寒武纪早期已存在明确的浅水环境下后生动物参与热带浅水碳酸盐生产工厂建造的实例[56],且苗岭世浅水环境后生动物参与工厂建造较为活跃[57],因而研究区浅水碳酸盐沉积环境中后生动物的匮乏与寒武纪生命演化过程并无直接关联。需要注意的是,寒武纪能够参与浅水生物礁建造的后生动物主要为古杯和其他海绵类等,这些后生动物骨骼壳体较薄且多孔,波浪和风暴条件下易被打碎、破坏,难以适应强水动力条件[25⁃26]。研究区微生物碳酸盐生产工厂发育期风暴盛行,可见大量的砾屑、鲕粒和风暴打碎的微生物岩角砾堆积(图4,5)。微生物群落主导形成的大型柱状、近球状和树枝状结构具有明显格架和黏结结构,能够抵御较强水动力条件的影响(图5)。另一方面,风暴盛行的环境条件带来大量营养盐,并且通过交互作用将正常浪基面之下富有机质和活性元素(磷、铁、锌等)水体带入潮坪环境[50];营养水平的上升有利于微生物群落的繁盛,为工厂建造提供了有利的营养条件[58]。对崮山组沉积相研究的新进展[40]显示,当时华北整体处于海侵过程,浅水环境微生物岩记录并不丰富,分布也较为零星。以河南云梦山地区为代表的古丈期微生物碳酸盐生产工厂发育,从现有证据来看仅具区域属性。由于研究区与现代潮坪环境鲨鱼湾相对局限的Hamelin Pool具有很大的相似性,我们推测两者水体盐度和饱和度等条件较为相似,即研究区微生物碳酸盐生产工厂亦处于碳酸盐矿物过饱和、盐度较高的局限环境。这可能是窄盐性后生动物难以参与研究区工厂建造的因素之一。
研究区微生物碳酸盐生产工厂的最终关闭可能与苗岭世古丈晚期淹没事件有关。单一类型碳酸盐生产工厂的消失一般与区域或全球范围内气候、环境条件的突然改变存在关联。例如,寒武纪第二世第三期华南汉南—米仓山地区热带浅水碳酸盐生产工厂的消亡与当时气候由干旱向潮湿转变造成陆源输入通量的显著提高有关[56]。二叠纪—三叠纪之交全球热带浅水碳酸盐生产工厂的崩溃及随后鲕粒工厂和灰泥工厂的兴盛,与全球范围生物大绝灭及随后的极端高温事件有关[59]。研究区微生物碳酸盐生产工厂虽然样式丰富但单期厚度非常有限(海底至水面高度一般不足1 m),具有很大的环境局限性,因而突然的海水加深对潮坪环境微生物碳酸盐生产工厂的运转带来致命性打击(图5c),工厂类型也会发生转换(如灰泥工厂)。从研究区岩石记录来看,崮山组上段基本为薄层泥晶条带灰岩和中厚层砾屑灰岩,代表古丈期云梦山地区整体海平面处于不断加深的趋势(直到末期才水体变浅),这也与当时华北板块和全球海平面变化趋势总体一致[60⁃61]。
-
(1) 研究区寒武纪苗岭世古丈期微生物碳酸盐生产工厂以底栖微生物群落为主要建造者,样式包括叠层石、凝块石、树枝石、均一石,以及MISS构造。单期工厂发育规模较小但空间延伸广,包括半球状、穹窿状、板状等宏观形态,结构上存在单独的建造样式,更多的是以复合形式构成的垂向组合。
(2) 微生物碳酸盐生产工厂发育基底为潮下带具平台状、塔状泥晶灰岩。基底常具被侵蚀形态,周围保存大量竹叶状砾屑以及异地搬运鲕粒形成的混杂堆积,这些特征指示工厂形成于风暴间歇期。单期工厂整体形成于潮坪环境变浅序列,中止时常被灰泥沉积(泥晶灰岩)掩盖。
(3) 微生物碳酸盐生产工厂发育与营养水平关系密切。研究区风暴盛行的气候背景下,通过强烈的海水搅动和再悬浮作用,正常浪基面之下的有机质和活性元素(磷、铁、锌等)被输入至潮坪,显著提高了当地营养水平。微生物碳酸盐生产工厂发育中止与快速海侵造成工厂被淹没有关,而工厂最终关闭与当时整体海侵背景下研究区适宜微生物碳酸盐大规模发育的环境因素(咸化、动荡、富营养)缺失有关。
(4)提供了一个与现代澳大利亚鲨鱼湾Hamelin Pool微生物碳酸盐生产工厂类似的古代实例,通过定量化评价方法对工厂内部样式、类型和构成进行了精细解剖,为认识深时碳酸盐生产工厂发育和消亡过程提供了一种新的思路。
Composition, Characteristics, and Sedimentological Significance of the Microbial Carbonate Factory in Deep Time: A case study from the Yunmengshan section, Henan province, China
-
摘要: 目的 “碳酸盐(生产)工厂”研究强调生物群落对碳酸盐生产过程以及规模的调控和特异性影响,在深入认识碳酸盐岩成因和形成机制方面具有重要价值,是碳酸盐沉积学领域关注热点和重要方向。虽然该概念的正式提出已有二十多年历史,但如何从工厂角度解读地质历史时期,特别是深时特异性、大规模碳酸盐岩的发育过程和影响因素方面仍缺乏系统性认识。 方法 以河南云梦山剖面寒武系崮山组为例,利用多尺度、网格定量化评价方法对微生物碳酸盐生产工厂的样式、内部结构、建造过程等方面进行了详细阐述。 结果 研究区微生物碳酸盐生产工厂以底栖微生物群落为主要建造者,形成样式包括叠层石、凝块石、树枝石、均一石以及微生物诱导沉积构造(MISS)。伴生的颗粒类型包括鲕粒、球粒、生屑、竹叶状内碎屑和微生物岩角砾,主要分布于工厂建造格架间。微生物碳酸盐生产工厂发育具期次性,单期工厂发育规模较小(海底至水面高度不足1 m)但空间延伸广,包括半球状、穹窿状、板状等宏观形态;其内部结构包括少量单独的建造样式,以及广泛发育的垂向复合形式。 结论 通过分析影响工厂发育的各类因素,局限潮坪环境下较高的营养水平对微生物碳酸盐生产工厂的繁盛可能起到了主要作用,而古丈期总体海侵背景造成适宜微生物碳酸盐生产工厂发育的环境消失,导致工厂最终关闭。相关工作的开展提供了一个与现代澳大利亚Hamelin Pool微生物碳酸盐生产工厂非常相似的古代实例,对认识寒武纪碳酸盐生产工厂类型、特征和演化规律方面具有较好的参考价值。Abstract: Objective The study of the "Carbonate Factory" emphasizes the specific effects of biological communities on the carbonate production process and scale moderation, contributing to our understanding of large-scale carbonate sedimentation. Although the concept and application of the "Carbonate Factory" have gained attention in carbonate sedimentology, there is still a lack of systematic understanding of how to interpret the origin, developmental processes, and influencing factors of specific types of carbonate sedimentation in geological history, particularly in deep time. Methods This study investigated the Gushan Formation of the Cambrian Miaolingian in the Yunmengshan area (Henan Province) as an example of a microbial carbonate factory. A multi-scale grid-based quantitative evaluation method was used to describe the specific form, internal structure, and construction process of the carbonate factory. Results Our results show that benthic microbial communities were the primary builders of the microbial carbonate factory in the study area. They formed unique structures, including the stromatolite, thrombolite, leiolite, and microbial-induced sedimentary structure (MISS). Various particle types, including ooids, flat pebbles, and microbialite breccias, were preserved within the microbial buildups. The development of the microbial carbonate factories occurred episodically during the Guzhangian Age (Miaolingian) at relatively small scales (height of <1 m); however, they extended widely in space and exhibited different macroscopic shapes such as subspherical, domical, and tabular. The internal structures of the microbial carbonate factories were made up of individual forms and vertical combinations of composite forms. Conclusions By analyzing various factors influencing the development of these factories, we found that higher nutrient levels may have played a crucial role in their development in the tide-flat environment. Additionally, the flooding event during the Guzhangian led to the eventual closure of these factories. This study provides an ancient example, similar to the modern microbial carbonate factory of the Hamelin Pool in Australia, which is a good reference for understanding the types, characteristics and evolutionary patterns of Cambrian carbonate factories.
-
Key words:
- microbialite /
- carbonate sedimentation process /
- nutritional conditions /
- Guzhangian /
- North China Block
-
图 1 华北寒武纪苗岭世古地理图及研究区岩石地层单元划分
Figure 1. Palaeogeography of the North China Block in the Cambrian Miaolingian, and the lithostratigraphic column of the study area

图 3 研究区崮山组微生物碳酸盐生产工厂岩石学特征
Figure 3. Photographs showing the lithological characteristics of the microbial carbonate factory preserved in the Gushan Formation
图 4 研究区典型微生物碳酸盐生产工厂复合结构类型和主要特征
Figure 4. Field photographs of the composite structures preserved within the microbial carbonate factory, with corresponding sketch maps and grid⁃based statistics
-
[1] Pomar L. Carbonate systems[M]//Scarselli N, Jurgen A, Chiarella D, et al. Regional geology and tectonics: Principles of geologic analysis. 2nd ed. Netherlands: Elsevier, 2020: 235-311. [2] 颜佳新, 孟琦, 王夏, 等. 碳酸盐工厂与浅水碳酸盐岩台地:研究进展与展望[J]. 古地理学报,2019,21(2):232-253. Yan Jiaxin, Meng Qi, Wang Xia, et al. Carbonate factory and carbonate platform: Progress and prospects[J]. Journal of Palaeogeography (Chinese Edition), 2019, 21(2): 232-253. [3] Pomar L, Haq B U. Decoding depositional sequences in carbonate systems: Concepts vs experience[J]. Global and Planetary Change, 2016, 146: 190-225. [4] Wilson J L. Carbonate facies in geologic history[M]. New York: Springer, 2012: 1-417. [5] James N P. Facies models 7. Introduction to carbonate facies models[J]. Geoscience Canada, 1977, 4(3): 123-125. [6] Betzler C, Brachert T C, Nebelsick J H. The warm temperate carbonate province: A review of the facies, zonations, and delimitations[J]. Courier Forschungsinstitut Senckenberg, 1997, 201(201): 83-99. [7] Halfar J, Godinez-Orta L, Mutti M, et al. Nutrient and temperature controls on modern carbonate production: An example from the gulf of California, Mexico[J]. Geology, 2004, 32(3): 213-216. [8] Michel J, Borgomano J, Reijmer J J G. Heterozoan carbonates: When, where and why? A synthesis on parameters controlling carbonate production and occurrences[J]. Earth-Science Reviews, 2018, 182(1): 50-67. [9] 李飞, 武思琴, 刘柯. 鲕粒原生矿物识别及对海水化学成分变化的指示意义[J]. 沉积学报, 2015, 33(3): 500-511. Li Fei, Wu Siqin, Liu Ke. Identification of ooid primary mineralogy: A clue for understanding the variation in paleo-oceanic chemistry[J]. Acta Sedimentologica Sinica, 2015, 33(3): 500-511. [10] Opdyke B N, Wilkinson B H. Carbonate mineral saturation state and cratonic limestone accumulation[J]. American Journal of Science, 1993, 293(3): 217-234. [11] Pomar L, Hallock P. Carbonate factories: A conundrum in sedimentary geology[J]. Earth-Science Reviews, 2008, 87(3/4): 134-169. [12] Schlager W. Carbonate sedimentology and sequence stratigraphy[M]. Tulsa: SEPM, 2005: 1-208. [13] Schlager W. Sedimentation rates and growth potential of tropical, cool-water and mud-mound carbonate systems[M]//Insalaco E, Skelton P W, Palmer T J. Carbonate platform systems: Components and interactions. London: Geological Society, London, Special Publications, 2000, 178(1): 217-227. [14] Schlager W. Benthic carbonate factories of the Phanerozoic[J]. International Journal of Earth Sciences, 2003, 92(4): 445-464. [15] Allwood A C, Walter M R, Kamber B S, et al. Stromatolite reef from the Early Archaean era of Australia[J]. Nature, 2006, 441(7094): 714-718. [16] Burne R V, Moore L S. Microbialites: Organosedimentary deposits of benthic microbial communities[J]. Palaios, 1987, 2(3): 241-254. [17] Grey K, Awramik S M. Handbook for the study and description of microbialites[M]. East Perth: Geological Survey of Western Australia, 2020: 1-278. [18] Suosaari E P, Reid R P, Mercadier C, et al. The microbial carbonate factory of Hamelin Pool, Shark Bay, Western Australia[J]. Scientific Reports, 2022, 12(1): 12902. [19] Riding R. Microbial carbonates: The geological record of calcified bacterial-algal mats and biofilms[J]. Sedimentology, 2000, 47(Suppl.1): 179-214. [20] Li X, Li F, Wang J G, et al. Formation and preservation of Eocene lacustrine microbialites in the western Qaidam Basin (northeastern Qinghai-Tibetan Plateau, China): Petrological, mineralogical, and geochemical constraints[J]. Sedimentary Geology, 2022, 440: 106257. [21] Feldmann M, McKenzie J A. Stromatolite-thrombolite associations in a modern environment, Lee Stocking Island, Bahamas[J]. Palaios, 1998, 13(2): 201-212. [22] Jahnert R J, Collins L B. Characteristics, distribution and morphogenesis of subtidal microbial systems in Shark Bay, Australia[J]. Marine Geology, 2012, 303-306: 115-136. [23] Grotzinger J P, Knoll A H. Stromatolites in Precambrian carbonates: Evolutionary mileposts or environmental dipsticks?[J]. Annual Review of Earth and Planetary Sciences, 1999, 27: 313-358. [24] Grotzinger J P, James N P. Precambrian carbonates: Evolution of understanding[M]//Grotzinger J P, James N P. Carbonate sedimentation and diagenesis in the evolving Precambrian world. Tulsa: SEPM Special Publication, 2000: 3-20. [25] 李雅兰, 李飞, 吕月健, 等. 陕南勉县寒武系仙女洞组生物礁岩相学及古环境分析[J]. 沉积学报, 2024,42(2):608-618. Li Yalan, Li Fei, Yuejian Lü, et al. Petrographic features and paleoenvironmental significance of the Lower Cambrian reef in the Xiannüdong Formation, Mian county, southern Shaanxi[J]. Acta Sedimentologica Sinica, 2024, 42(2): 608-618. [26] 李杨凡, 李飞. 前寒武—寒武纪重大转折期生物礁是如何演化的?[J]. 地球科学, 2022, 47(10):3853-3855. Li Yangfan, Li Fei. How did reefs evolve during the Precambrian-Cambrian transition?[J]. Earth Science, 2022, 47(10): 3853-3855. [27] Li F, Deng J T, Kershaw S, et al. Microbialite development through the Ediacaran-Cambrian transition in China: Distribution, characteristics, and paleoceanographic implications[J]. Global and Planetary Change, 2021, 205: 103586. [28] 邓嘉婷, 李飞, 龚峤林, 等. 埃迪卡拉纪—寒武纪之交微生物岩特征对比及古海洋学意义:以汉南—米仓山地区为例[J]. 古地理学报, 2021, 23(5): 919-936. Deng Jiating, Li Fei, Gong Qiaolin, et al. Characteristics and palaeoceanographic significances of microbialite development in the Ediacaran-Cambrian transition: A case study from Hannan-Micangshan area[J]. Journal of Palaeogeography (Chinese Edition), 2021, 23(5): 919-936. [29] Chen Z Q, Tu C Y, Pei Y, et al. Biosedimentological features of major microbe-metazoan transitions (MMTs) from Precambrian to Cenozoic[J]. Earth-Science Reviews, 2019, 189: 21-50. [30] Kershaw S, Crasquin S, Li Y, et al. Microbialites and global environmental change across the Permian-Triassic boundary: A synthesis[J]. Geobiology, 2012, 10(1): 25-47. [31] Collins L B, Jahnert R J. Stromatolite research in the Shark Bay world heritage area[J]. Journal of the Royal Society of Western Australia, 2014, 97(1): 189-219. [32] Papineau D, Walker J J, Mojzsis S J, et al. Composition and structure of microbial communities from stromatolites of Hamelin Pool in Shark Bay, Western Australia[J]. Applied and Environmental Microbiology, 2005, 71(8): 4822-4832. [33] Logan B W, Davies G R, Read J F, et al. Carbonate sedimentation and environments, Shark Bay, Western Australia[M]. Tulsa: AAPG, 1970: 1-205. [34] Tang H, Kershaw S, Liu H, et al. Permian-Triassic boundary microbialites (PTBMs) in Southwest China: Implications for paleoenvironment reconstruction[J]. Facies, 2017, 63(1): 1-23. [35] Tang H, Kershaw S, Tan X C, et al. Sedimentology of reefal buildups of the Xiannüdong Formation (Cambrian Series 2), SW China[J]. Journal of Palaeogeography, 2019, 8: 11. [36] Mata S A, Bottjer D J. Microbes and mass extinctions: Paleoenvironmental distribution of microbialites during times of biotic crisis[J]. Geobiology, 2012, 10(1): 3-24. [37] Wan B, Tang Q, Pang K, et al. Repositioning the Great Unconformity at the southeastern margin of the North China Craton[J]. Precambrian Research, 2019, 324: 1-17. [38] 牟传龙, 周恳恳, 陈小炜, 等. 中国岩相古地理图集(埃迪卡拉纪—志留纪)[M]. 北京: 地质出版社, 2016: 1-154. Mou Chuanlong, Zhou Kenken, Chen Xiaowei, et al. Atlas of lithofacies and paleogeography in China, Ediacaran-Silurian[M]. Beijing: Geological Publishing House, 2016: 1-154. [39] 刘印环, 王建平, 张海清, 等. 河南的寒武系和奥陶系[M].北京: 地质出版社, 1991: 1-254. Liu Yinhuan, Wang Jianping, Zhang Haiqing,et al. The Cambrian and Ordovician systems of Henan province [M]. Beijing: Geological Publishing House, 1991: 1-254. [40] Xin H, Chen J T, Gao B, et al. Spatio-temporal distribution of the Cambrian maceriate reefs across the North China Platform[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2023, 614: 111429. [41] 裴放, 张海清, 阎国顺, 等. 河南省地层古生物研究(第三分册)早古生代(华北型)[M]. 郑州: 黄河水利出版社, 2008: 1-302. Pei Fang, Zhang Haiqing, Yan Guoshun, et al. The Third Paleozoic study of stratigraphic paleontology in Henan province (Early Paleozoic)[M]. Zhengzhou: The Yellow River Water Conservancy Press, 2008: 1-302. [42] Shapiro R S. A comment on the systematic confusion of thrombolites[J]. Palaios, 2000, 15(2): 166-169. [43] Michel J, Laugié M, Pohl A, et al. Marine carbonate factories: A global model of carbonate platform distribution[J]. International Journal of Earth Sciences, 2019, 108(6): 1773-1792. [44] 李飞, 易楚恒, 李红,等. 微生物成因鲕粒研究进展[J]. 沉积学报, 2022, 40(2): 319-334. Li Fei, Yi Chuheng, Li Hong, et al. Recent advances in ooid microbial origin: A review[J]. Acta Sedimentologica Sinica, 2022, 40(2): 319-334. [45] Jiang L Q, Feely R A, Carter B R, et al. Climatological distribution of aragonite saturation state in the global oceans[J]. Global Biogeochemical Cycles, 2015, 29(10): 1656-1673. [46] Flügel E. Microfacies of carbonate rocks: Analysis, interpretation and application[M]. Berlin: Springer, 2010: 1-984. [47] Millero F J, Lee K, Roche M. Distribution of alkalinity in the surface waters of the major oceans[J]. Marine Chemistry, 1998, 60(1/2): 111-130. [48] Arp G, Reimer A, Reitner J. Microbialite formation in seawater of increased alkalinity, Satonda Crater Lake, Indonesia[J]. Journal of Sedimentary Research, 2003, 73(1): 105-127. [49] Schultze L K P, Merckelbach L M, Carpenter J R. Storm-induced turbulence alters shelf sea vertical fluxes[J]. Limnology and Oceanography Letters, 2020, 5(3): 264-270. [50] Rumyantseva A, Lucas N, Rippeth T, et al. Ocean nutrient pathways associated with the passage of a storm[J]. Global Biogeochemical Cycles, 2015, 29(8): 1179-1189. [51] Hoover R S, Hoover D, Miller M, et al. Zooplankton response to storm runoff in a tropical estuary: Bottom-up and top-down controls[J]. Marine Ecology Progress Series, 2006, 318: 187-201. [52] Mutti M, Hallock P. Carbonate systems along nutrient and temperature gradients: Some sedimentological and geochemical constraints[J]. International Journal of Earth Sciences, 2003, 92(4): 465-475. [53] Hallock P. Coral reefs, carbonate sediments, nutrients, and global change[M]//Stanley G D. The history and sedimentology of ancient reef systems. Boston: Springer, 2001: 387-427. [54] Hallock P, Schlager W. Nutrient excess and the demise of coral reefs and carbonate platforms[J]. Palaios, 1986, 1(4): 389-398. [55] Stanley S M, Hardie L A. Secular oscillations in the carbonate mineralogy of reef-building and sediment-producing organisms driven by tectonically forced shifts in seawater chemistry[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1998, 144(1/2): 3-19. [56] Li H, Li F, Li X, et al. Development and collapse of the Early Cambrian shallow-water carbonate factories in the Hannan-Micangshan area, South China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2021, 583: 110665. [57] Lee J H, Riding R. Marine oxygenation, lithistid sponges, and the early history of Paleozoic skeletal reefs[J]. Earth-Science Reviews, 2018, 181: 98-121. [58] 李杨凡, 李飞, 王夏, 等. 上扬子北缘寒武纪早期后生动物礁特征及古环境意义[J]. 地球科学,2023,48(11):4321-4334. Li Yangfan, Li Fei, Wang Xia, et al. Sedimentary characteristics and paleoenvironmental significance of Early Cambrian metazoan reefs in northern margin of Upper Yangtze Block[J]. Journal of Earth Science, 2023,48(11):4321-4334. [59] Li F, Gong Q L, Burne R V, et al. Ooid factories operating under hothouse conditions in the earliest Triassic of South China[J]. Global and Planetary Change, 2019, 172: 336-354. [60] Haq B U, Schutter S R. A chronology of Paleozoic sea-level changes[J]. Science, 2008, 322(5898): 64-68. [61] Chen J T, Chough S K, Han Z Z, et al. An extensive erosion surface of a strongly deformed limestone bed in the Gushan and Chaomidian Formations (late Middle Cambrian to Furongian), Shandong province, China: Sequence-stratigraphic implications[J]. Sedimentary Geology, 2011, 233: 129-149. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 899
- HTML全文浏览量: 213
- PDF下载量: 156
- 被引次数: 0

