

 下载:
下载:


-
早侏罗世是泛大陆(Pangea)裂解关键时期,并伴随着三叠纪—侏罗纪界线中大西洋岩浆岩省(Central Atlantic Magmatic Province,CAMP)和早侏罗世托阿尔早期卡鲁—费拉尔大火成岩省(Karoo-Ferrar Large Igneous Province,Karoo-Ferrar LIP)的相继喷发。在两次大火成岩省喷发的背景下,早侏罗世是以周期性全球气候环境剧烈变化、海洋缺氧以及频繁生物危机为标志的时期。三叠纪末生物大灭绝事件(End Triassic Extinction,ETE)和早侏罗世托阿尔早期大洋缺氧事件(Toarcian Oceanic Anoxic Event,T-OAE)是地质历史时期重要的极热事件,因此其特征、成因以及生物和环境变化已经得到了大量的研究[1⁃3]。近年来,越来越多的学者发现了早侏罗世其他重要的碳同位素扰动事件,诸如辛涅缪尔期—普林斯巴期界线事件(Sinemurian-Pliensbachian Boundary Event,SPBE)、margaritatus菊石带事件(margaritatus zone event,ME)、margaritatus-spinatum菊石带界线事件(margaritatus-spinatum boundary event,MSBE)和普林斯巴期—托阿尔期界线事件(Pliensbachian-Toarcian Boundary Event,PTBE)[4⁃10]。然而,相比于ETE和T-OAE,目前这些事件受到关注较少,因此相关的气候—环境变化研究仍然非常薄弱。
普林斯巴期的碳同位素扰动事件主要是以碳同位素负偏移为特征的SPBE、MSBE和PTBE,和以碳同位素正偏移为特征的ME[4,7⁃12]。目前为止,这些事件较经典的记录来自Mochras钻孔(英国)[8]、Breggia剖面(瑞士)[9]、卧龙剖面(藏南)[10]、Sancerre-Couy钻孔(法国)[12]以及Amellago-IKis剖面(摩洛哥)[13]。近年来的研究更深入地探讨了普林斯巴期的气候环境变化,如普林斯巴期区域和全球范围内气候变暖和变冷[4,7,14⁃16],以及区域和全球海平面的波动[14,17⁃20]。值得注意的是,ME之前的辛涅缪尔期—普林斯巴期过渡时期,全球海相和陆相有机质和无机碳酸盐的δ13C均记录了一次剧烈的碳同位素负偏移事件,被称为辛涅缪尔期—普林斯巴期界线事件(SPBE)[6,9⁃10,13,21⁃22]。这个事件被认为可能与CAMP晚期喷发有关,而这一过程释放大量CO2注入大气—海洋系统,进而可能导致全球升温和大洋含氧量降低以及碳酸盐台地的生产力危机[9⁃10,13,22⁃23]。目前的研究表明,普林斯巴晚期ME很可能改善了SPBE时期以来恶化的生态环境,进而促进Lithiotis Fauna(造礁双壳)的扩散[5,10]。这些双壳是早侏罗世特有的造礁生物[24⁃26],直至T-OAE开始时期才彻底灭绝,可能与水体富营养化以及海洋酸化有关[5,27⁃28]。然而,由于剖面地层不完整、碳同位素曲线趋势不明显以及台地陆源碎屑输入等影响[5,10],ME和MSBE对普林斯巴晚期—托阿尔(最早)期Lithiotis Fauna繁盛和碳酸盐台地在全球广泛发育可能产生的影响仍然不明确。
本研究提供了早侏罗世时期位于南半球特提斯喜马拉雅西段的藏南札达地区东秋剖面的普林斯巴晚期—托阿尔(最早)期的高分辨率有孔虫生物地层、碳同位素地层、微相特征以及生屑变化等数据。利用上述资料与特提斯喜马拉雅中段定日地区和西特提斯进行对比,进而探讨该时期碳循环扰动以及相应的气候环境变化对该地区的碳酸盐台地演化过程可能影响。
-
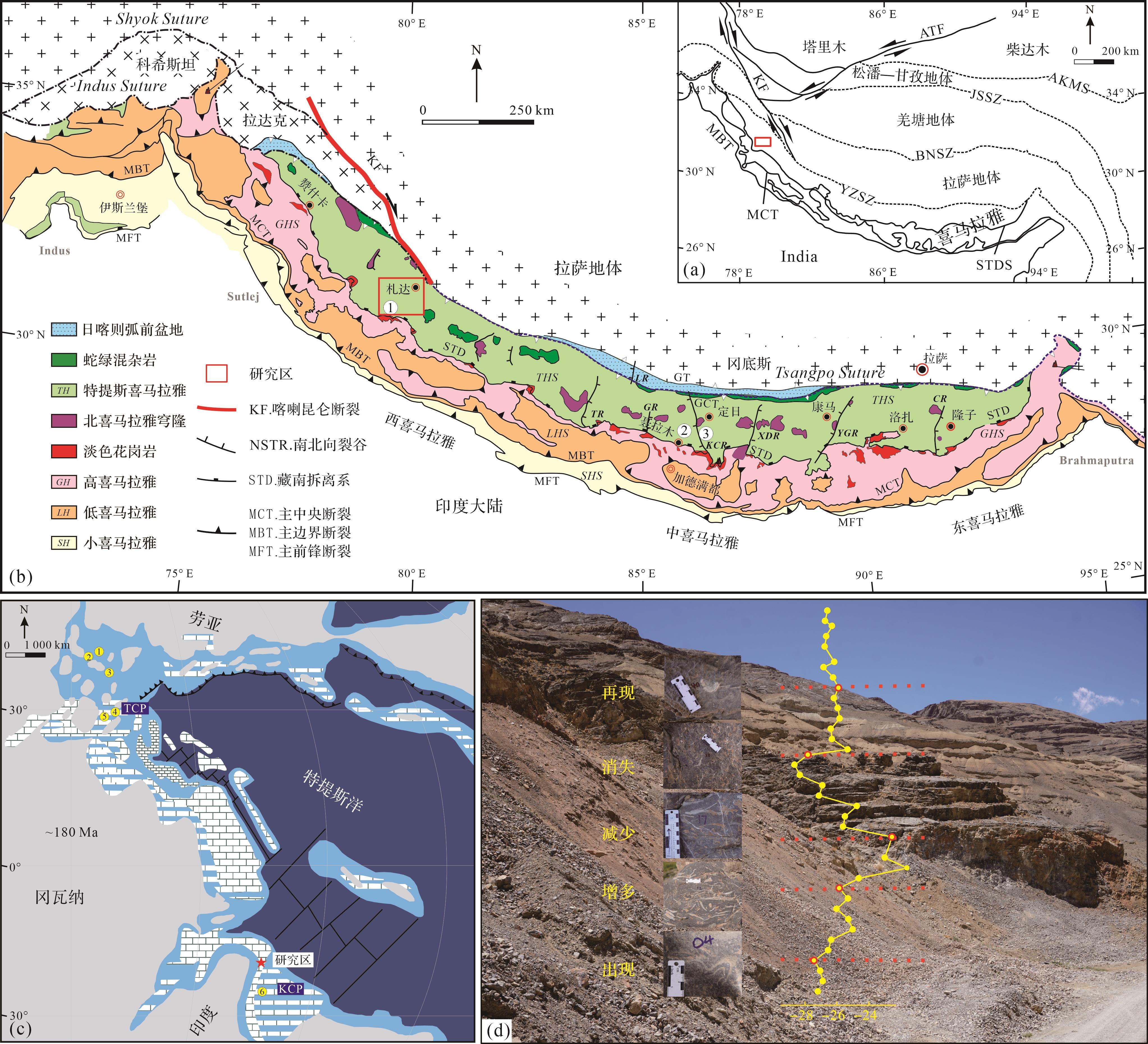
喜马拉雅造山带是由印度—欧亚大陆碰撞所形成的陆—陆碰撞造山带,其岩石组合被主前缘断裂(MFT)、主边界断裂(MBT)、主中央断裂(MCT)和藏南拆离系(STDS)分为四部分:由南至北分别为小喜马拉雅、低喜马拉雅、高喜马拉雅和特提斯喜马拉雅(图1a,b)[29⁃31]。其中,特提斯喜马拉雅带代表印度大陆北缘沉积(图1c),分为南北两个亚带,其南亚带主要是以混合的浅水碳酸盐岩和陆架陆源碎屑岩沉积为特征,而北亚带是以深水沉积为特征[32⁃35]。在早侏罗世,特提斯喜马拉雅南亚带从赞什卡(印度)至我国西藏南部广泛发育碳酸盐岩沉积,被称为Kioto碳酸盐台地(图1c)[32⁃34,36⁃37]。

图 1 特提斯喜马拉雅构造背景和早侏罗世研究区地质背景
Figure 1. Tethys Himalayan tectonic setting and the Early Jurassic geological setting of the study area
特提斯喜马拉雅普林斯巴期地层被命名为普普嘎组。在特提斯喜马拉雅中段定日和聂拉木地区,普普嘎组主要为灰白色中—厚层生屑颗粒灰岩和鲕粒颗粒灰岩的浅水台地沉积,富含造礁双壳Lithiotis Fauna[20,34],这些特征和特提斯喜马拉雅西段札达地区有很好的可对比性[38⁃39]。普普嘎组的下部和上部分别发现了Lithiotis Fauna(造礁双壳)和Orbitopsellapraecursor大型底栖有孔虫,表明其属于普林斯巴期;普普嘎组和聂聂雄拉组过渡带为托阿尔大洋缺氧事件(TOAE)的层位,指示该组最上部属于托阿尔最早期[10,20,33⁃34,40⁃41]。因此,普普嘎组的时代为普林斯巴期—托阿尔(最早)期。
-
东秋剖面在藏南札达县(31°28'59.59" N,78°38'50.51" E),位于特提斯喜马拉雅西南部,其地层属于普普嘎组。在东秋剖面采集54个碳酸盐岩样品并将每个样品制成岩石薄片,用徕卡DM1000显微镜观察岩石薄片并鉴定有孔虫。有孔虫鉴定和分类主要参考Loeblich et al.[42]和Boudagher-Fadel[43⁃44]的分类体系。本研究依据大型底栖有孔虫(Larger Benthic Foraminifera,LBF)进行生物地层LBF带划分,并将其与特提斯喜马拉雅中段定日地区和西特提斯LBF带进行对比,以确定其准确的沉积时代。
-
采用点计法定量统计54张岩石薄片碳酸盐颗粒的含量。依据Riding[45]的划分方案,将待统计组分划分为基质(M)、胶结物(C)以及颗粒(S)三大类,将其中的颗粒进一步划分为碳酸盐颗粒(鲕粒、团块、内碎屑、核形石、似球粒、生物碎屑和晶粒)和陆源碎屑颗粒,并做成Excel表作为PhotoQuad[46]软件定名模板。利用PhotoQuad软件将薄片图像圈出一定大小(1.8 cm×2.7 cm)的合适区域并等间距投点,按照Excel表对每个点逐一定名。结果随点计法统计点数的增加而变得更准确[47],因此对每张薄片统计375个点,进而确保统计结果的准确性。
特别说明的是,颗粒的保存状态对统计结果有一定影响,但整体上不会影响最终的统计结果[48]。根据沉积构造、结构以及定量统计颗粒和生物化石组成与含量,可以确定每张薄片的微相类型。在浅水碳酸盐台地环境中,碳酸盐生屑颗粒的含量和多样性对海平面和环境的变化非常敏感[49]。利用碳酸盐生屑颗粒相对含量变化的定量数据,建立了Kioto碳酸盐台地更详细的沉积演化特征。同时,本研究将与特提斯喜马拉雅中段定日地区和西特提斯Trento碳酸盐台地(意大利)的相关研究进行对比,进而更全面地了解早侏罗世普林斯巴晚期—托阿尔(最早)期特提斯碳酸盐台地生物演化过程[5⁃6,10,20]。
-
在东秋剖面共选取34个样品进行全岩无机碳氧同位素分析。在获得粉末过程中,避开胶结物充填的裂隙和孔隙或较大的生物碎屑,尽量确保所获取的数据反映原始沉积信号。样品粉末在70 ℃下烘烤去湿,然后用磷酸(H3PO4)溶解,生成的CO2在氦气的输送下进入质谱仪进行测量。全岩无机碳氧同位素是在中国地质调查局成都地质调查中心所完成,所用仪器为Finnigan MAT 253稳定同位素质谱仪。无机碳同位素结果是以Vienna Peedee Belemnite(VPDB)为参照标准。δ13Ccarb和δ18Ocarb的分析精度为±0.06‰和±0.08‰。
选取40个样品进行有机碳同位素分析。操作步骤如下:在约5 g碳酸盐岩粉末中加入过量2 mol/L的盐酸去除无机碳酸盐岩,离心后加入去离子水再离心,倒掉上清液,去除残余的HCl,重复上述清洗过程直至上清液pH值为中性;称取约1.5 mg烘干样品包入锡杯中进行测试。有机质碳同位素是在中国地质调查局成都地质调查中心所完成,所用仪器为Finnigan MAT 253稳定同位素质谱仪。参照标准为国产标样碳黑GBW04407(δ13CVPDB=-22.43‰±0.07‰)。仪器对δ13Corg分析精度为±0.1‰。
-
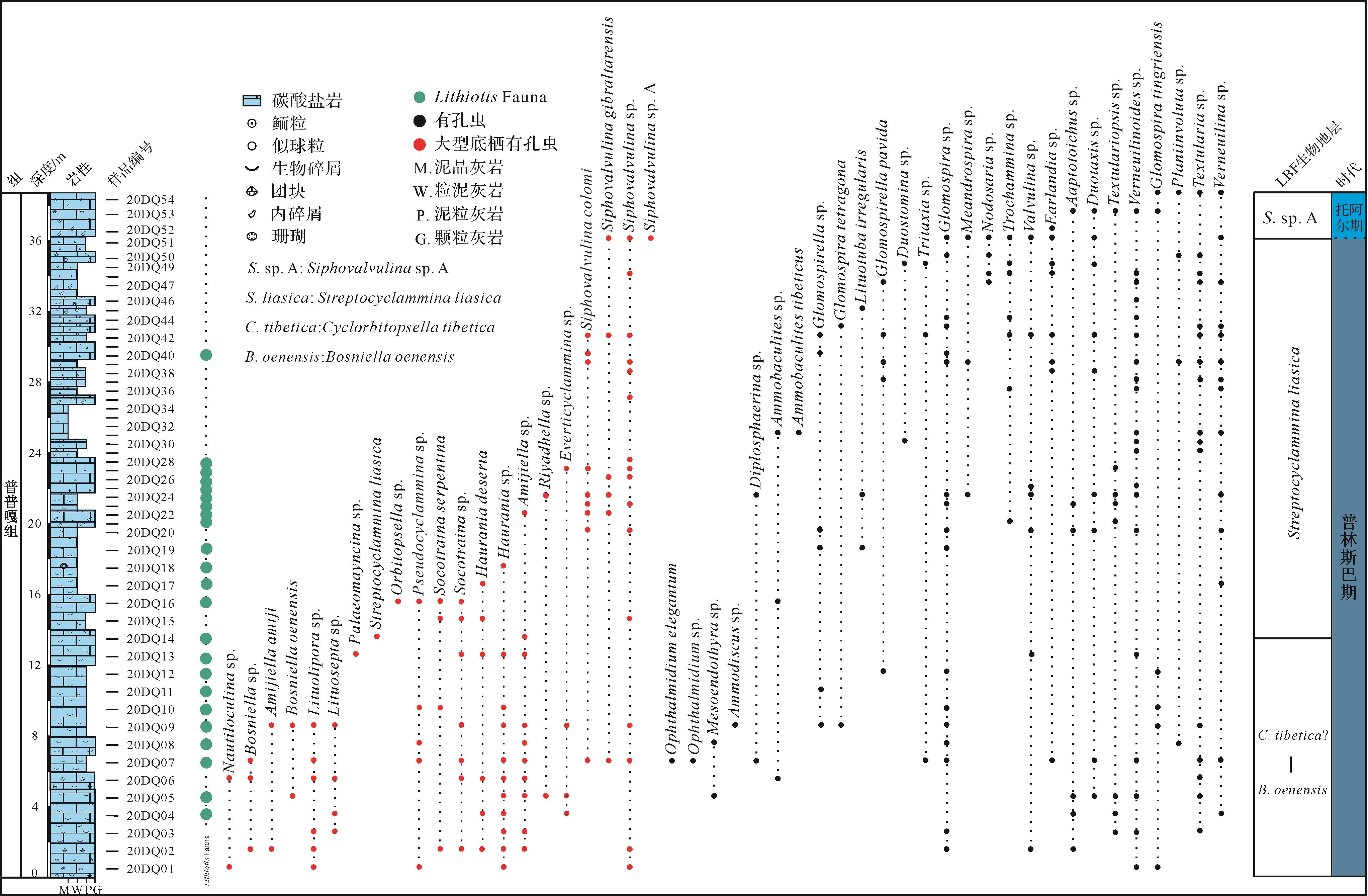
对在东秋剖面所采集的54块样品进行有孔虫鉴定,识别出有孔虫化石414个,涉及35属48种。在底栖有孔虫中,有一组内部结构复杂、尺寸相对较大的物种被称为大型底栖有孔虫(larger benthic foraminifera,LBF)。根据东秋剖面大型底栖有孔虫(LBF)地层分布情况,并结合有孔虫特征及其多样性,本研究将剖面的有孔虫分布自下而上划分为三个有孔虫带(图2),具体结果如下。

图 2 特提斯喜马拉雅西段札达县东秋剖面岩性柱,有孔虫分布和生物地层学框架
Figure 2. Lithological log, foraminiferal distribution, and biostratigraphical framework in the Dongqiu section of Zanda county, western segment of the tethys Himalaya
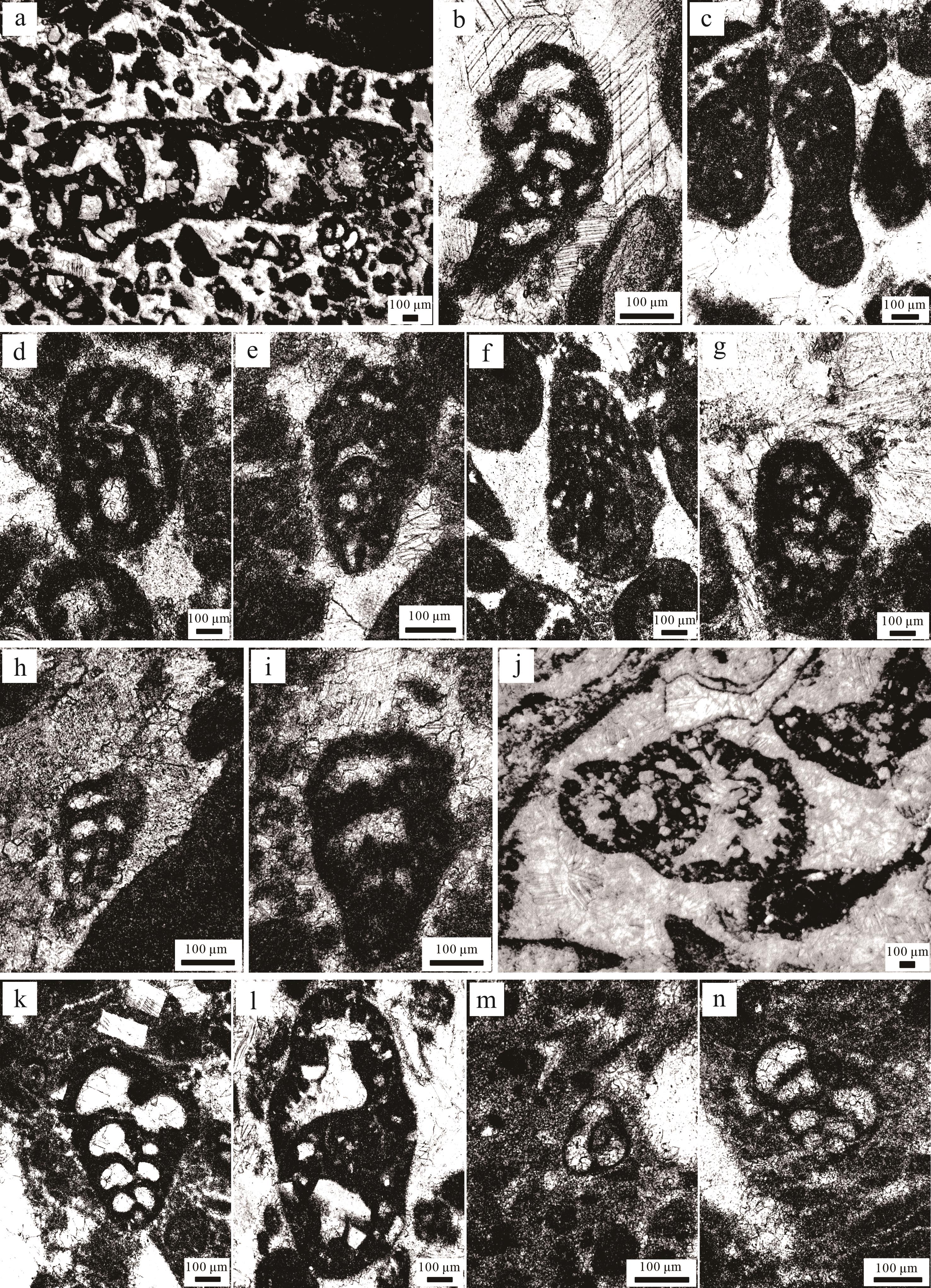
(1) Bosniella oenensis-Cyclorbitopsella tibetica(?)带位于东秋剖面下部(0~13.5 m,这一范围为近似值,下同),其底界以Bosniella oenensis(图3a)的首现定义,顶界为Streptocyclammina liasica的首现位置[10]。Bosniella oenensis在西特提斯的辛涅缪尔阶上部和普林斯巴阶下部普遍存在[44]。然而,最近的研究表明特提斯喜马拉雅中段定日地区卧龙剖面的该有孔虫出现在上普林斯巴阶的下部地层中[10]。东秋剖面中,普普嘎组缺失上普林斯巴阶下部的Cyclorbitopsella tibetica带[10]。本带LBF主要包括Bosniella oenensis、Bosniella sp.(图3b)、Lituosepta sp.(图3c)、Lituolipora sp.(图3d)、Haurania sp.(图3e)、Socotraina sp.(图3f)、Pseudocyclammina sp.(图3g)、Siphovalvulina sp.(图3h)以及Amijiella sp.(图3i)。Mesoendothyra sp.、Textularia sp.、Verneuilinoides sp.等有孔虫在该带也比较常见。

图 3 札达县东秋剖面普林斯巴晚期—托阿尔(最早)期有孔虫组合图
Figure 3. Late Pliensbachian⁃Earliest Toarcian foraminiferal assemblage in the Dongqiu section of Zanda county
(2) Streptocyclammina liasica带位于东秋剖面中上部(13.5~36 m),其底界以Streptocyclammina liasica(图3j)的首现定义,顶界为Siphovalvulina sp.A的首现位置[10]。在西特提斯和东特提斯喜马拉雅中段定日地区,Streptocyclammina liasica从普林斯巴阶上部到托阿尔阶下部普遍存在[10,44]。该带LBF主要包括Streptocyclammina liasica、Siphovalvulina sp.、Siphova lvulina colomi(图3k)以及Everticyclammina sp.(图3l);有孔虫在该带上部出现得较多,例如Textularia sp.、Verneuilinoides sp.、Earlandia sp.、Trochammina sp.以及Glomospira sp.等。
(3) Siphovalvulina sp. A带位于东秋剖面顶部(36~38.5 m),其底界以Siphovalvulina sp. A(图3m)的首现定义,未见顶界[10]。该带与东特提斯喜马拉雅中段定日地区卧龙剖面LBF带是一致的,但是西特提斯对应的是Socotraina serpentina带[44]。该带LBF主要包括Siphovalvulina sp. A、Siphovalvulina sp.以及Siphovalvulina gibraltarensis(图3n)。Earlandia sp.、Glomospira sp.、Duotaxis sp.以及Glomospiratingriensis等有孔虫在该带经常出现。
特提斯喜马拉雅西段东秋剖面LBF带与中段定日地区卧龙剖面LBF带基本一致,但是对比卧龙剖面,东秋剖面缺失Cyclorbitopsella tibetica生物带[10]。这些LBF带整体上可与西特提斯同期碳酸盐台地的LBF带相当,尤其是摩洛哥、意大利以及西班牙南部记录的LBF带[44,50⁃51]。有孔虫带表明东秋剖面主体属于上普林斯巴阶,顶部可能延伸至托阿尔阶最底部,这和该剖面大量出现的Lithiotis所指示的时代是一致的[5,10]。根据碳同位素地层,特提斯喜马拉雅中段定日地区卧龙剖面SPBE延伸至Bosniella oenensis带顶部[10],而东秋剖面Bosniella oenensis带并未发现碳同位素负偏移的特征。综上所述,东秋剖面的时代属于普林斯巴晚期—托阿尔(最早)期。
-
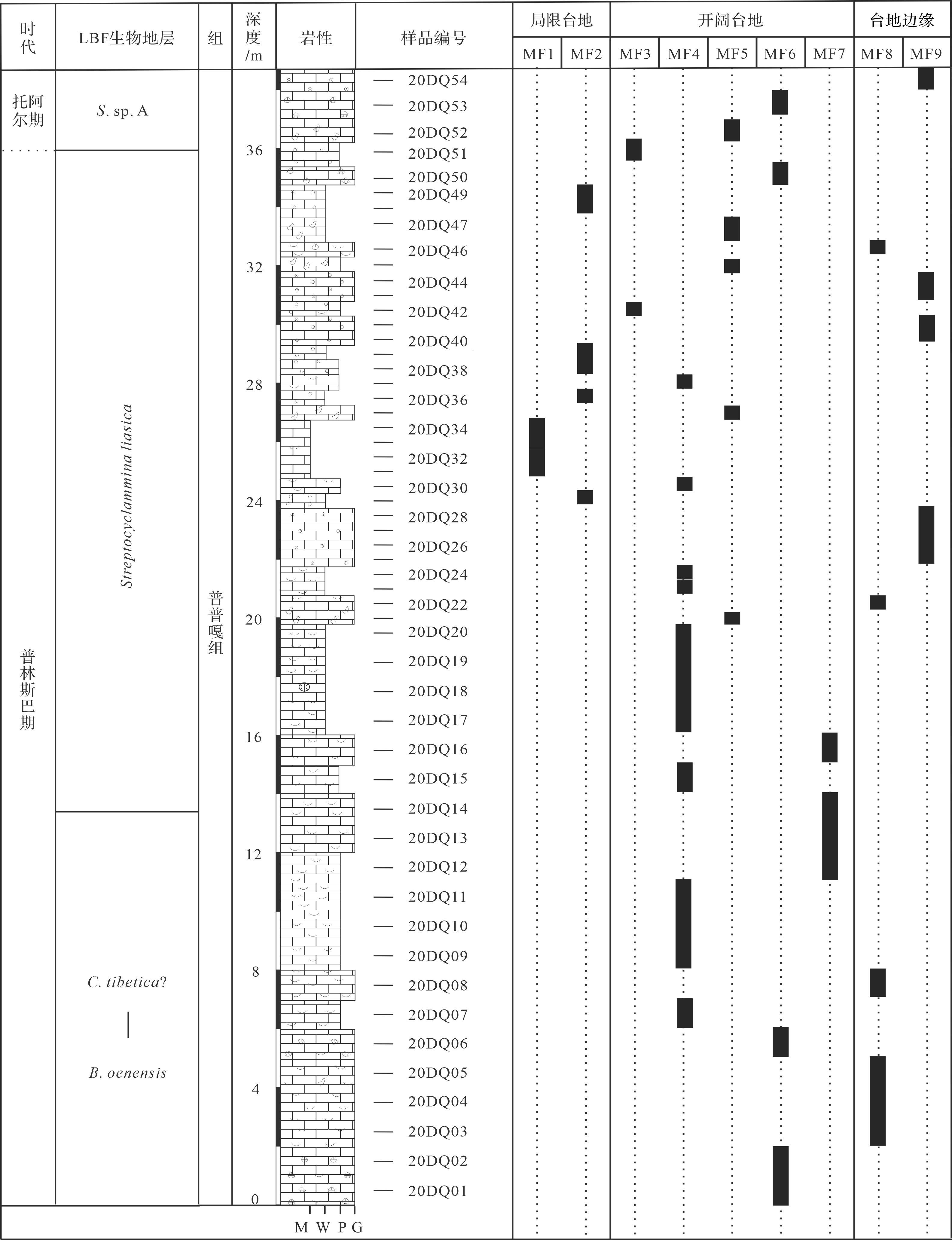
根据沉积构造、结构和定量统计颗粒和生物化石组成与含量,在东秋剖面共识别出9种碳酸盐岩微相(图4,5),分别是泥晶灰岩(MF1)、白云石化似球粒粒泥灰岩(MF2)、生屑似球粒泥粒灰岩(MF3)、生屑泥粒/粒泥灰岩(MF4)、内碎屑泥粒—颗粒灰岩(MF5)、团块颗粒灰岩(MF6)、Lithiotis砾屑灰岩(MF7)、生屑颗粒灰岩(MF8)和鲕粒颗粒灰岩(MF9)。沉积微相和微相组合表明特提斯喜马拉雅西段在普林斯巴晚期—托阿尔(最早)期发育浅水碳酸盐台地环境,包括局限台地(MF1~MF2)、开阔台地(MF3~MF7)以及台地边缘(MF8~MF9)等。

图 4 札达县东秋剖面普普嘎组地层和微相分布
Figure 4. Stratigraphy and microfacies distribution in the Pupuga Formation of the Dongqiu section, Zanda county

图 5 札达县东秋剖面典型微相类型和野外Lithiotis Fauna变化
Figure 5. Typical microfacies types and Lithiotis Fauna changes in the Dongqiu section of Zanda county
-
1) MF1:泥晶灰岩
以泥晶方解石基质为主,生屑颗粒含量为0~8.8%,平均值为4.3%,仅局部可见薄壳双壳(图5a)。泥晶基质含量高和生物碎屑的含量极少表明其沉积于能量低、水循环局限、盐度异常的局限台地环境[49]。MF1出现在普普嘎组中上部,但较少见(图4)。
2) MF2:白云石化似球粒粒泥灰岩
颗粒主要是细粒似球粒,含量为8.5%~28.3%,平均值为14.6%,分选磨圆度好,分布不均匀。生屑含量为3.7%~11.2%,平均值为5.6%,可见双壳、钙质海绵骨针以及腹足等,局部可见少量内碎屑、生物潜穴以及纹层状构造(图5b)。该微相存在局部白云岩化作用,多数白云石为自形—半自形菱形并具有环带状结构。纹层状构造在潮上带和潮间带较为常见[49]。生物种类和含量少以及颗粒之间被泥晶基质所充填表明该微相沉积环境盐度异常,能量较低。纹层状构造以及较弱的白云岩化作用表明沉积环境最可能为潮间带中上部[49]。MF2常出现在普普嘎组中上部(图4)。
-
1) MF3:生屑似球粒泥粒灰岩
颗粒主要是细粒似球粒和生屑,颗粒含量大于50%(图5c)。似球粒含量为34.7%~35.2%,平均值为34.9%,分选性较好,为椭圆状或是次圆状至圆状。生屑含量为15.5%~19.8%,平均值为17.6%,主要为钙质海绵骨针、薄壳双壳,也见有孔虫、腹足以及介形虫等正常底栖生物。颗粒之间被泥晶基质所充填。生物种类以及数量中等以及颗粒之间被泥晶基质所充填表明其沉积环境最可能为低—中等强度水循环、水动力低的开阔台地[49]。MF3出现在普普嘎组上部,但较少见(图4)。
2) MF4:生屑粒泥/泥粒灰岩
颗粒主要为生物碎屑,偶见内碎屑、似球粒(图5d)。生物碎屑种类丰富,含量为8.3%~52.8%,平均值为26.7%,主要为双壳碎片、有孔虫、腕足、腹足以及珊瑚(图5j)等正常底栖生物,偶尔可见少量钙质海绵骨针、海百合、薄壳双壳、Lithiotis Fauna(其含量小于10%)等。其中绝大多数生物内部结构是保存完好的,但大量双壳可能是被波浪打碎成双壳碎片。颗粒之间被泥晶基质充填。生物种类多、含量丰富以及泥晶基质中高能颗粒等特征指示沉积环境最可能位于具有开阔水体循环的较低能滩间,高能的碳酸盐颗粒可以在波浪的作用下搬运至此[49]。MF4出现在普普嘎组中部(图4)。
3) MF5:内碎屑泥粒灰岩—颗粒灰岩
颗粒主要是内碎屑,含量为31.7%~54.7%,平均值为37.8%,分选中等,大部分为砂屑,多为椭圆状或是次圆状至圆状,表层呈褐色氧化圈(图5e)。次要颗粒为生物碎屑,含量为9.3%~17.1%,平均值为14.3%,常见双壳、腹足、腕足、海百合以及有孔虫等,多被泥晶套包裹。颗粒之间多为亮晶方解石,部分为泥晶基质。颗粒之间充填物表明其受到较强水动力影响,但不足以将泥晶完全冲洗干净。颗粒支撑结构以及泥晶化作用强烈的生物碎屑等特征指示其最可能沉积于开阔台地中具有较强水动力作用的环境[49]。MF5常出现在普普嘎组中上部(图4)。
4) MF6:团块颗粒灰岩
颗粒主要为团块、生物碎屑以及鲕粒,颗粒含量大于70%(图5f)。团块含量为29.6%~45.9%,平均值为39.9%。团块经过微生物黏结作用而形成的,一般较为圆滑,也有黏结成不规则状;其内部无构造,一般有多个不规则“核心”。生物碎屑含量为12.5%~26.1%,平均值为20.0%,种类较为丰富,较常见双壳、有孔虫、腹足、腕足以及棘皮类动物等,泥晶化作用强烈,表面多裹有泥晶套。颗粒间主要为亮晶方解石,局部有泥晶充填。填隙物含有少量泥晶表明其未能受到持续稳定的水动力影响。团块、丰富的泥晶化强烈生屑以及少部分鲕粒表明其最可能位于离颗粒滩较近,具有正常水体循环且能量较强的开阔台地[49]。MF4常出现在普普嘎组底部和顶部(图4)。
5) MF7:Lithiotis砾屑灰岩
颗粒主要为生物碎屑、内碎屑、似球粒,颗粒含量大于50%。生屑颗粒含量为45.3%~57.1%,平均值为51.2%,其种类丰富,常见Lithiotis Fauna(图5g)、其他较小双壳、钙藻、有孔虫以及海百合等。在生屑颗粒中,Lithiotis Fauna(造礁双壳)约占70%,在薄片中展示的长度为0.5~1.0 cm,在野外记录中长度为2~15 cm(图5k~p),因此我们将其定名为Lithiotis砾屑灰岩。Lithiotis Fauna作为造礁生物,常生活在鲕粒滩背后的浅水地区[23,52]。较高的颗粒含量、颗粒支撑、生屑含量较高以及特有的Lithiotis Fauna生物指示其最可能位于开阔台地与台地边缘过渡处,有正常水体循环的沉积环境[49]。MF7常出现在普普嘎组下部(图4)。
-
1) MF8:生屑颗粒灰岩
颗粒主要为生物碎屑、团块、内碎屑、似球粒,含量大于70%。生屑颗粒含量为25.8%~44.2%,平均值为33.9%,种类丰富,常见双壳、有孔虫、腹足、海百合、腕足、藻类和苔藓虫等,泥晶化作用非常强烈,大多生物包裹泥晶套(图5h)。颗粒之间为亮晶方解石胶结物,与大多数海百合共轴生长。生屑种类丰富、含量高、强烈泥晶化作用和颗粒支撑均表明其沉积环境位于具有开阔水体循环的台地边缘高能生屑滩[49]。MF8常出现在普普嘎组底—中部(图4)。
2) MF9:鲕粒颗粒灰岩
颗粒主要是鲕粒,含量为40.3%~62.7%,平均值为46.8%,直径为0.2~0.4 mm,分选性好,磨圆度较好,其核心多为粒度不等的似球粒颗粒,有时为生屑颗粒。绝大多数鲕粒为放射—同心鲕(图5i),少部分鲕粒发生强烈泥晶化作用,圈层结构遭到破坏,结构模糊,部分也有鲕粒相互黏结为团块。生屑颗粒含量为1.9%~17.3%,平均值为11.0%,可见孔虫、双壳、腹足等。颗粒之间为镶嵌状亮晶方解石胶结物,为大气淡水渗流胶结。颗粒支撑结构、丰富的鲕粒、颗粒强烈泥晶化都指示台地边缘高能浅水鲕粒滩沉积[49]。MF9常出现在普普嘎组中上部(图4)。
沉积微相和微相组合揭示东秋剖面的普普嘎组是几乎不受陆源碎屑影响的浅水碳酸盐台地沉积,和特提斯喜马拉雅中段定日和聂拉木地区类似[20]。东秋剖面普普嘎组记录了由开阔台地和台地边缘逐渐过渡到局限台地,后又迅速演变到开阔台地和台地边缘的沉积序列。普普嘎组底—中部沉积环境是以有正常水体循环的开阔台地和台地边缘,主要发育生屑粒泥/泥粒灰岩(MF4)、团块颗粒灰岩(MF6)、生屑颗粒灰岩(MF8)和鲕粒颗粒灰岩(MF9)。向上,沉积环境逐渐演变为局限台地,主要发育泥晶灰岩和白云石化似球粒粒泥灰岩(MF1~MF2)。随后,沉积环境很快又进入开阔台地和台地边缘,主要发育内碎屑泥粒灰岩—颗粒灰岩(MF5)、生屑颗粒灰岩(MF8)和鲕粒颗粒灰岩(MF9),偶尔出现生屑似球粒泥粒灰岩(MF3)。
-
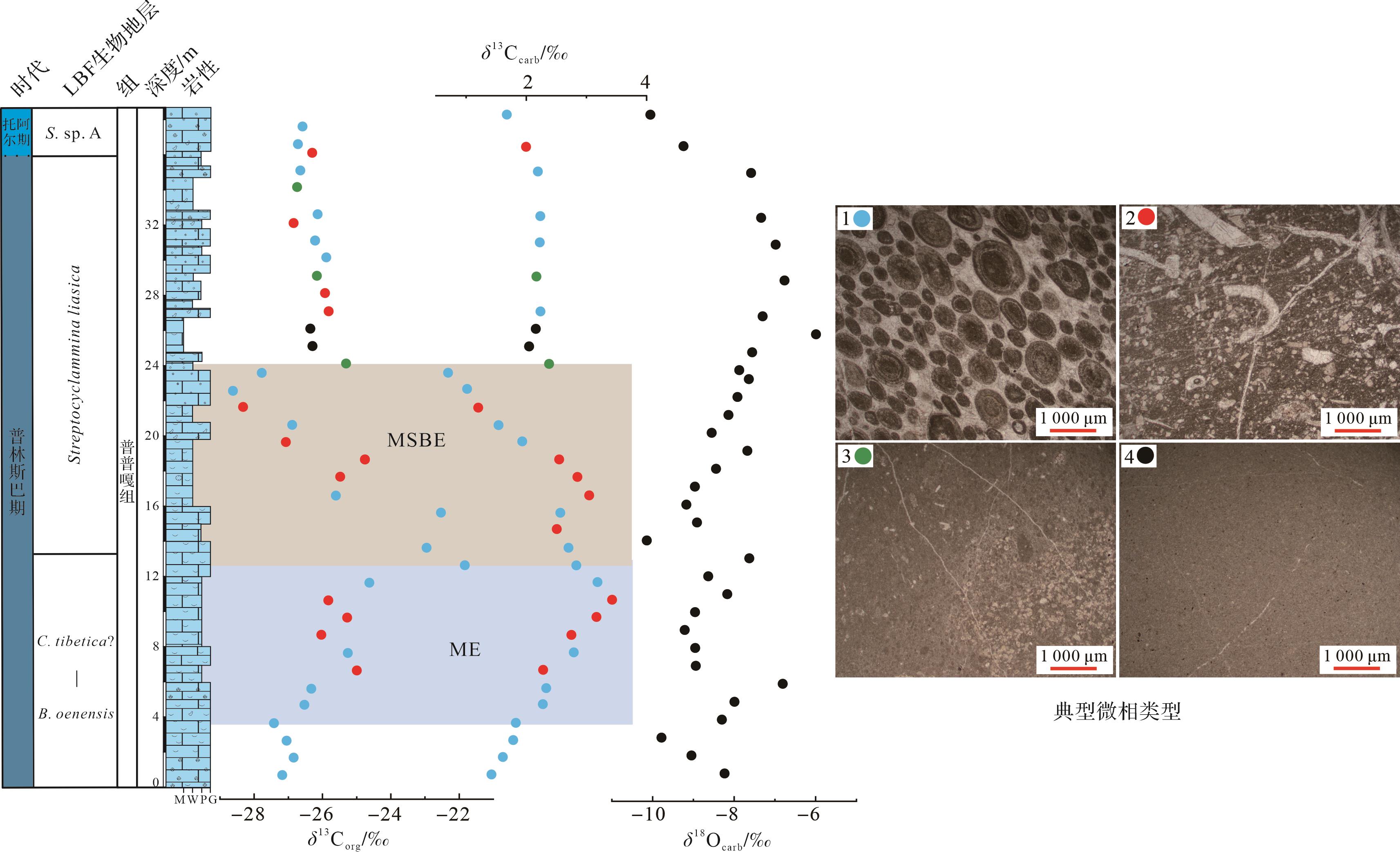
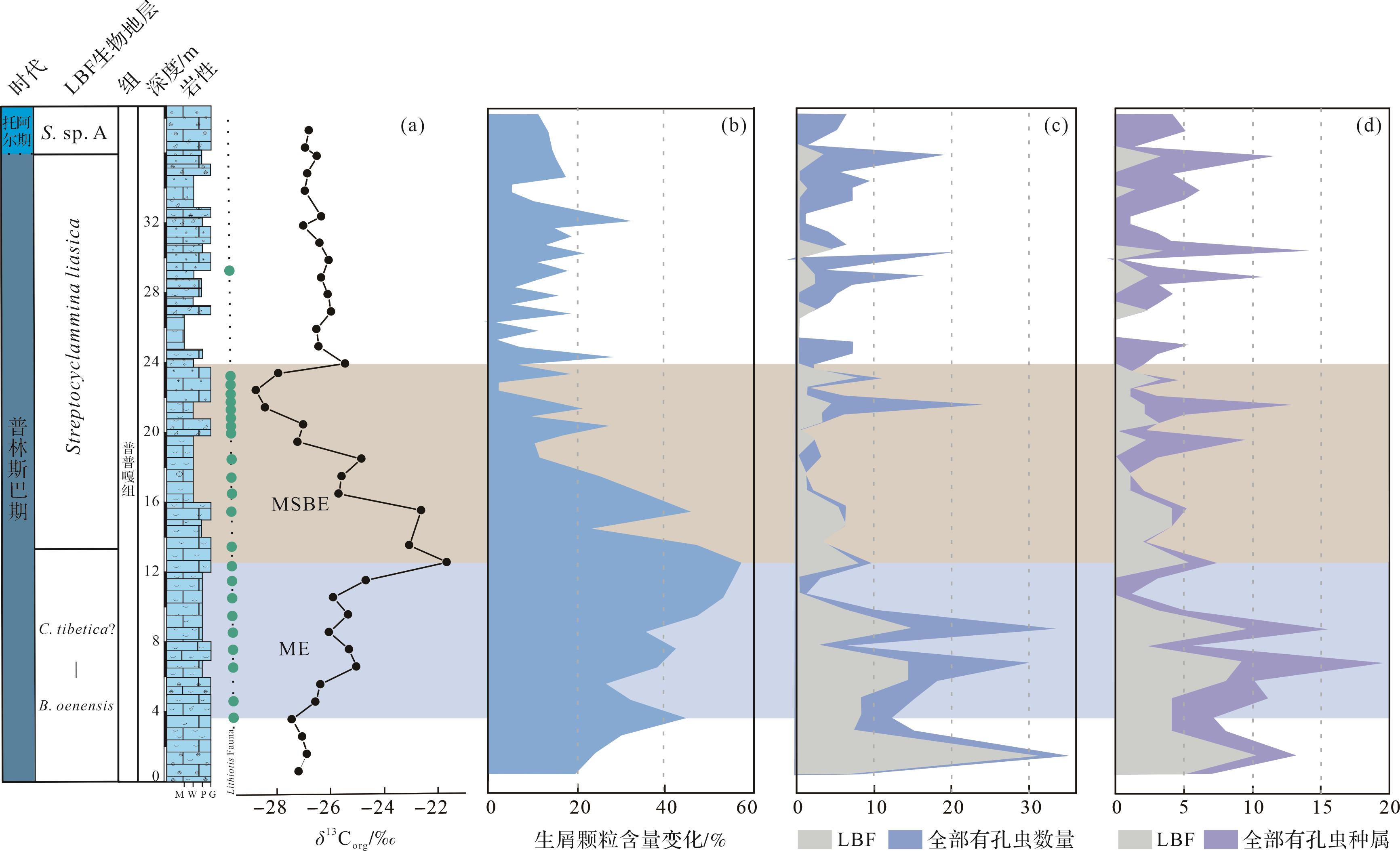
东秋剖面δ13Corg值介于-26.8‰~-21.6‰,平均值为-26.0‰。东秋剖面普普嘎组下部显示幅度为5‰~6‰的正偏移(3.5~12.5 m,图6),紧接着出现一个约6‰的负偏移(12.5~24 m,图6),在24 m之后,δ13Corg稳定在-26‰左右。δ13Ccarb值介于1.0‰~3.4‰,平均值为2.2‰。总体上,δ13Ccarb 与δ13Corg变化趋势基本一致,但是其偏移幅度较小,正负偏移幅度均约为1.5‰,负偏移之后,δ13Ccarb值在2‰左右变化。对应的氧同位素值(δ18Ocarb)介于-6.0‰~-11.6‰,平均值为-8.4‰。

图 6 札达县东秋剖面δ13Corg、全岩δ13Ccarb和δ18Ocarb
Figure 6. δ13Corg, bulk δ13Ccarb, and δ18Ocarb in the Dongqiu section of Zanda county
-
利用点计法定量统计生物碎屑颗粒的含量变化(图7b)。根据统计,生物碎屑颗粒变化范围为0~57.1%,平均值为20.6%。东秋剖面3.5~12.5 m段的生物碎屑主要有双壳、有孔虫、藻类、棘皮类、腕足、腹足、介形虫等,生物种类更为丰富,其含量变化范围为20.8%~57.1%,平均值为38.8%,在约12.5 m处达到高峰;在12.5~24 m段,生物碎屑主要是双壳、有孔虫,偶尔见棘皮类、腹足、珊瑚以及腕足,其含量变化范围为1.9%~47.2%,平均含量为18.9%,生物多样性和丰度较之前出现明显的下降。具体来说,生物碎屑颗粒含量从12.5~15.5 m开始缓慢下降,在15.5~23 m段开始快速下降,在21 m处降到相对低点为8.3%,在约22.5 m处降到最低点为1.9%。在约24 m之后,生屑颗粒其含量变化范围为0~29.3%,平均值为12.2%。具体来说,在24~27.5 m段,生屑颗粒含量仍处于较低的状态,其平均值为9.1%,在27.5 m之后生屑颗粒含量主要在15%左右波动。根据野外照片和薄片镜下特征,东秋剖面Lithiotis Fauna在约3.5 m处开始出现,此时其最大长度约为5.5 cm(图5k),而且在约9.5 m处,其数量和最大长度(约10 cm,图5l)开始明显地增加,在10.5~15.5 m段数量达到高峰(图5m)。在约16.5 m处其数量和最大长度(约7 cm,图5n)开始出现下降。在21.5~23.5 m段其数量和最大长度(约2 cm)达到最小值,表面风化较为严重(图5o),在镜下此段表现为鲕粒颗粒灰岩,且并未在薄片中发现这类双壳。在24~29 m,该生物完全消失不见。向上,Lithiotis Fauna在约29.5 m处再次出现,但其数量和最大长度(约2 cm,图5p)一直未达到开始下降之前的状态。

图 7 札达县东秋剖面生屑变化与碳同位素曲线的关系
Figure 7. Relationship between bioclastic change and carbon isotope curve in the Dongqiu section of Zanda county
由于大型底栖有孔虫(larger benthic foraminifera,LBF)对环境的变化响应更加敏锐,本文对有孔虫的数量以及种属进行统计,以便观察有孔虫的数量以及种属的变化情况(图7c,d)。通过薄片鉴定,我们共统计了48种有孔虫,共计414个,其中21种大型底栖有孔虫,共计170个。在0~12.5 m段,有孔虫的种属和数量保持较高水平,其平均值分别为8.4和13.4,其中LBF种属和数量也保持较高水平,其平均值分别为4.3和7.3。在12.5~24.0 m段,有孔虫的种属和数量呈现明显的下降趋势,其平均值分别为3.1和5.3,其中大型底栖有孔虫的种属和数量也呈现明显的下降趋势,其平均值分别为1.7和2.4。在这个区间内,10种大型底栖有孔虫消失,占比为66.7%(10/15)分别是Palaeomayncina sp.、Streptocyclammina liasica、Orbitopsella sp.、Pseudocycl ammina sp.、Socotraina serpentina、Socotraina sp.、Haurania deserta、Amijiella sp.以及Riyadhella sp.。在约24 m之后,有孔虫的种属和数量均值分别为3.8和5.6,其中LBF的种属和数量均值分别为0.5和0.6。
-
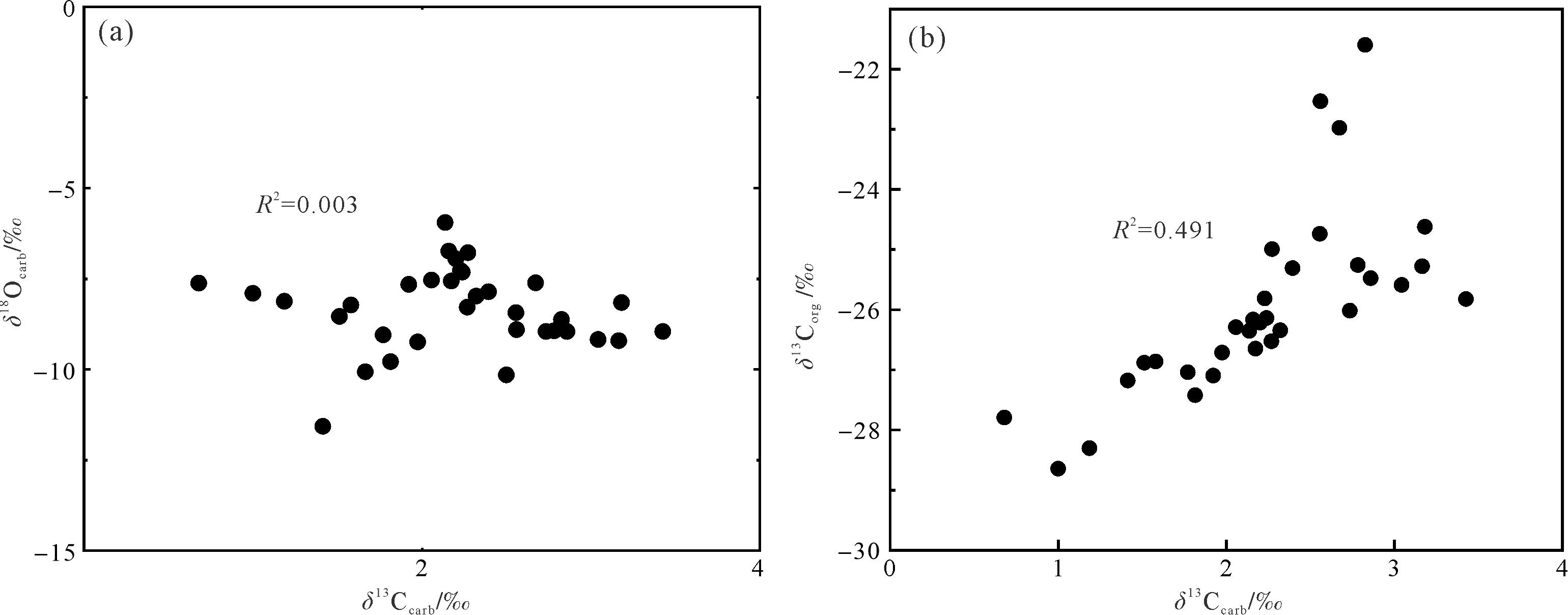
有机质来源的改变和成岩作用都可能对碳同位素产生较大的影响[53⁃54]。一般来说,δ13Ccarb和δ18Ocarb值之间存在相关性被认为受到成岩作用改造的影响,而不存在相关性则表明其未受成岩改造的影响[55⁃58]。研究中碳同位素来自浅水碳酸盐台地环境,其中δ18Ocarb和δ13Ccarb(R2=0.003)无相关性(图8a),表明δ13Ccarb受后期成岩作用较小。δ18O是成岩作用的敏感指标,其值小于-5‰通常被认为存在一定程度的成岩改造,在-5‰和-10‰之间则被认为成岩蚀变非常严重[59]。研究中绝大多数δ18Ocarb值小于-5‰,大于-10‰(图6),其平均值为-8.4‰,表明样品可能受到一定的成岩作用改造。另外,δ13Ccarb和δ13Corg之间存在耦合关系通常被认为是碳酸盐岩和有机质最初产生于海洋表面水域,并且它们保留了原始的δ13C成分,而δ13Ccarb和δ13Corg之间不存在耦合关系被认为受到成岩作用改造[60⁃63]。研究中δ13Corg和δ13Ccarb(R2=0.491)有较强相关性(图8b),表明δ13C保留了原始海水的信号。更重要的是:(1)研究中的δ13C的演化趋势也与同时期欧洲、非洲、北美以及中国藏南其他地区(定日和聂拉木)碳同位素具有良好的对比性[4,8⁃10,12⁃13,16];(2)δ13Ccarb值介于1.0‰~3.4‰,这与同时期东、西特提斯的δ13Ccarb值(1‰~3‰)一致[4⁃6,8,10,60]。综合所有证据表明,东秋剖面的δ13C保存较好,能较真实地反映早侏罗世普林斯巴晚期碳循环扰动的信号。

图 8 (a)δ13Ccarb⁃δ13Corg相关性图解;(b)δ13Ccarb⁃δ18Ocarb相关性图解
Figure 8. (a) Correlation between δ13Ccarb and δ13Corg; (b) correlation between δ13Ccarb and δ18Ocarb
岩性的变化也会影响δ13C的组成[53]。东秋剖面普普嘎组共有4个典型微相,分别是颗粒灰岩、泥粒/粒泥灰岩、白云石化似球粒粒泥灰岩和泥晶灰岩(图6)。从图6中可以看到,每一种微相类型的δ13Corg和δ13Ccarb值的分布范围都较广,表明研究区δ13C组成不受岩性变化的影响。
-
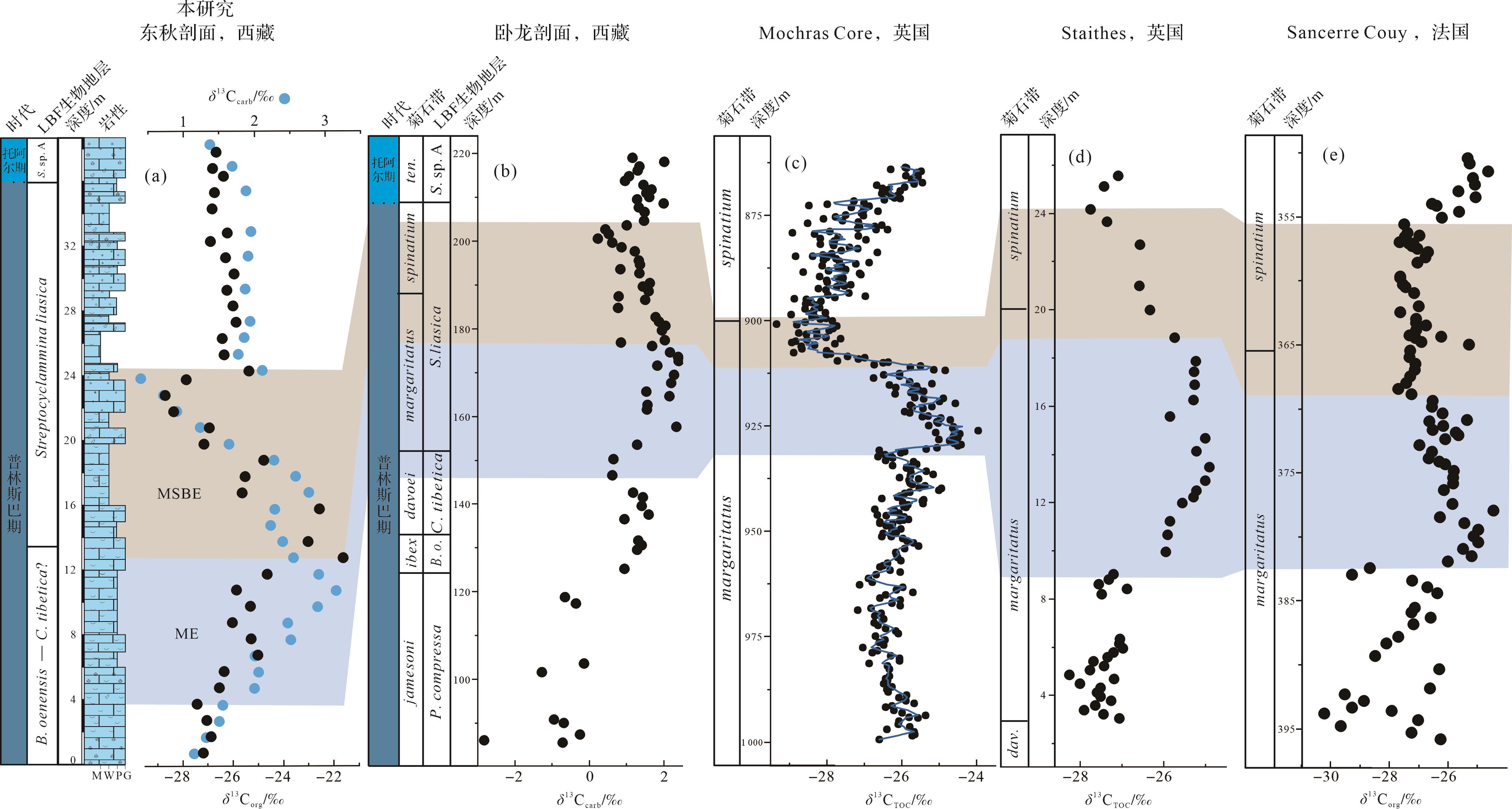
早侏罗世普林斯巴晚期发生多次碳同位素扰动事件,如margaritatus菊石带事件(ME)、margaritatus-spinatum菊石带界线事件(MSBE)和普林斯巴期—托阿尔期界线事件(PTBE)[5⁃11]。在有孔虫生物地层格架的基础上,将札达县东秋剖面普林斯巴晚期的碳同位素曲线与同处特提斯喜马拉雅的定日卧龙剖面以及西特提斯典型剖面进行对比(图9)。综合对比结果表明,东秋剖面普普嘎组B.oenensis-C.tibetica(?)带上部(3.5~12.5 m)观察到的δ13Ccarb(约1.5‰)和δ13Corg(5‰~6‰)正偏移应对应于卧龙剖面S.liasica带下部和欧洲(英国、法国)典型剖面margaritatus菊石带的碳同位素扰动事件(margaritatus zone event,ME)[8,10,12,16]。然而,从LBF带上来看,东秋剖面ME主要位于B.oenensis-C.tibetica(?)带上部(davoei菊石带),要稍早于卧龙剖面的S.liasica带下部(margaritatus菊石带),这可能是因为有孔虫带精度不够所导致的差异。其他地区海相和陆相同层位有机质和无机碳酸盐的δ13C也记录了这次正偏移,例如欧洲西班牙、葡萄牙以及瑞士等地区碳同位素记录了2‰~6‰的正偏移[4,8⁃9,12⁃14,64⁃66];北美美国俄勒冈州(Oregon)、阿拉斯加州(Alaska)和加拿大海达瓜伊群岛(Haida Gwaii)有机碳同位素记录了1.5‰~3‰的正偏移[67⁃68];北非阿尔及利亚(Algeria)和摩洛哥地区无机碳同位素记录了2‰~6‰的正偏移[7,13]。导致碳同位素正偏移的原因可能是该时期生产力增加,海洋浮游植物通过光合作用将更多的12C固定在有机质中,使得大量富含12C的有机质在海底埋藏[8,10,13,69⁃70]。目前的研究资料表明西班牙、葡萄牙和北美在该时期广泛出现富有机质的黑色页岩沉积[14,65,68,70]。值得注意的是在ME之前,早侏罗世辛涅缪尔期—普林斯巴期过渡时期发生一次全球碳同位素负偏移为特征的事件(SPBE),在全球海相和陆相同层位有机质和无机碳酸盐的δ13C都有响应[5⁃6,9⁃10,13,21⁃22]。根据同期硫同位素正偏移以及汞含量异常富集,这个事件可能与CAMP晚期喷发有关,释放大量CO2进入大气—海洋系统,进而导致全球升温和大洋含氧量降低[9,71]。随后,ME期间有机质的埋藏可能消耗了SPBE期间增加的CO2,进而导致阶段性变冷,这一现象被同时期其他剖面的箭石和腕足δ18O值的同步正偏移所证实[4,7,14⁃15,72]。

图 9 普林斯巴晚期碳同位素记录的全球对比
Figure 9. Global correlation of the carbon isotopic record of the Late Pliensbachian
在ME之后,东秋剖面普普嘎组S.liasica带下部(12.5~24.0 m)观察到的δ13Ccarb(约1.5‰)和δ13Corg(约6‰)负偏移应对应于卧龙剖面S.liasica带上部和欧洲典型剖面margaritatus-spinatum菊石带的碳同位素扰动事件(margaritatus-spinatum boundary event,MSBE)[4,8⁃10,12⁃13,16]。然而,从LBF带上来看,东秋剖面MSBE主要位于S.liasica带下部(margaritatus菊石带),要稍早于卧龙剖面的S.liasica带上部(margaritatus-spinatum菊石带),这同样可能是因为有孔虫带精度不够所导致的差异。在此期间,特提斯西部和北极大陆架地区的全岩碳酸盐岩以及腕足和箭石δ18O值最大、Mg/Ca值下降以及黏土矿物伊利石的增加和蒙脱石的减少,这些证据表明其可能是一个干冷时期[4,14,65,70,73⁃74]。在此期间,极地地区海平面下降、钙芒硝状方解石(glendonites)和冰碛岩的出现支持冰盖存在的假设[75⁃79],冰川控制的海退事件在西、东特提斯均有记录[10,14,19,80]。因此,该时期碳同位素负偏移被认为可能是全球可能存在海平面下降促使沉积物改造、有机物氧化和异化作用释放的大量轻碳而导致[4,13,65,70]。
普林斯巴期—托阿尔期界线碳同位素负偏移一般出现在MSBE之后,目前在欧洲、日本、北非摩洛哥甚至南美洲均有报道[11,81⁃86]。然而,东秋剖面并没有记录下与这一事件相关的碳同位素特征(图6,9),可能是因为海平面下降而引起的地层缺失。西特提斯和北极陆架海地区广泛报道了与全球变冷相关的海平面下降,时间上与普林斯巴期—托阿尔期界线事件(PTBE)一致[81,86⁃87]。在生物地层基础上,普林斯巴期—托阿尔期界线事件可能发生在东秋剖面普林斯巴阶上部33~36 m段。研究的东秋剖面普林斯巴阶上部(33~36 m)主要由内碎屑泥粒灰岩—颗粒灰岩(MF5)迅速转变到白云石化似球粒粒泥灰岩(MF2),表明此期间存在海平面下降。因此,研究认为可能是由于这次海平面下降引起地层缺失,从而使东秋剖面未记录普林斯巴期—托阿尔期界线事件。
-
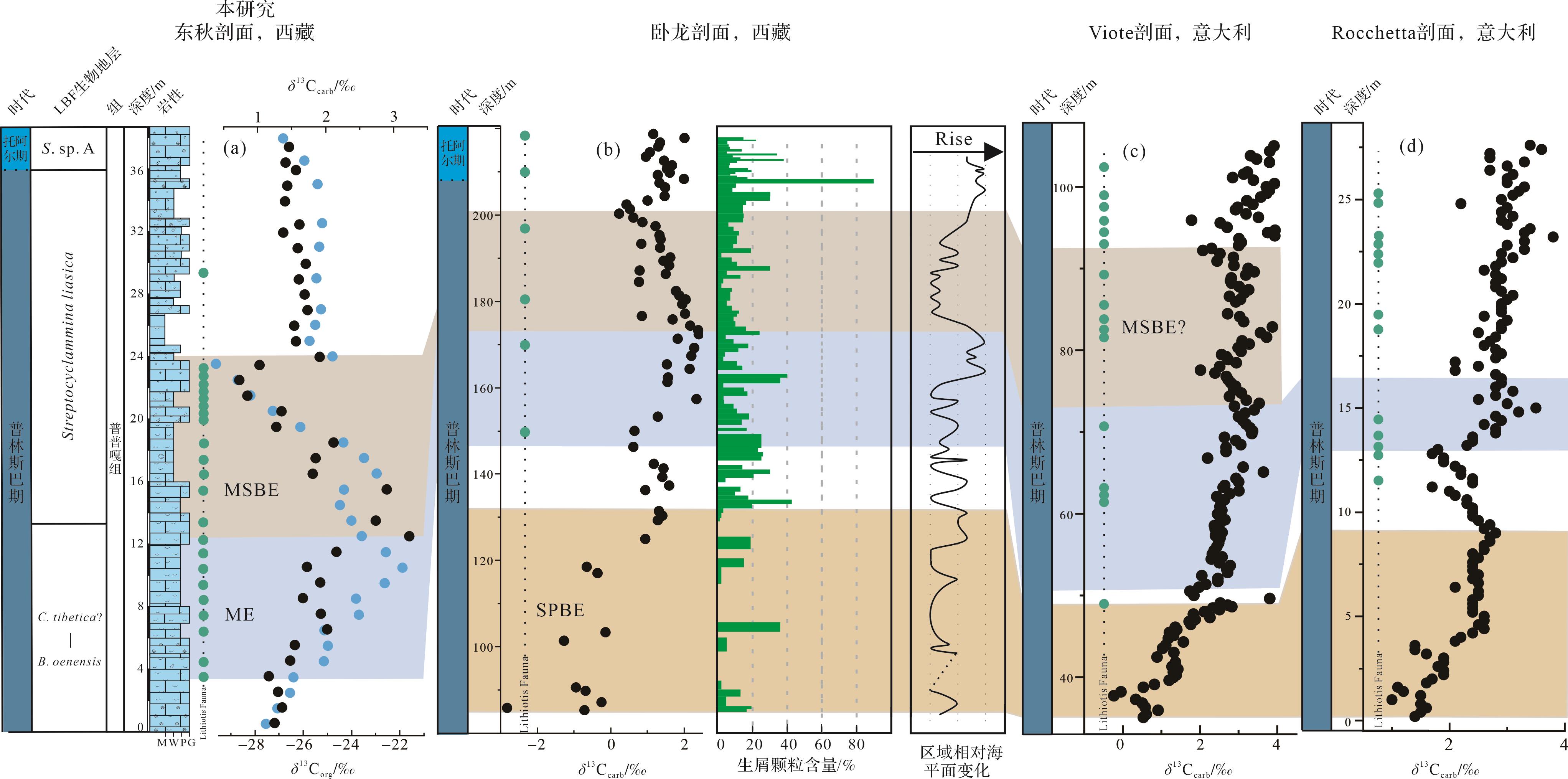
根据东秋剖面的微相变化、生屑颗粒含量变化、大型底栖有孔虫种属数量和Lithiotis Fauna分布与碳同位素事件之间的关系(图7),研究探讨了ME和MSBE对浅水碳酸盐台地生物和沉积环境的影响。在ME期间,生屑颗粒含量呈现增长的趋势以及LBF种属和数量处于较高的水平。特别地,Lithiotis Fauna(大型造礁双壳)在ME开始时出现(约3.5 m),并向上逐渐变多(图10)。这些双壳是生物礁的重要建造者,代表了早侏罗世特有的全球范围的生物钙化事件,是地质历史时期首例全球扩散的造礁双壳[24,52],同时和造礁珊瑚伴生。同样地,在特提斯喜马拉雅中段卧龙剖面和西特提斯Trento碳酸盐台地Viote和Rocchetta剖面,Lithiotis Fauna首次出现也大致对应于δ13Ccarb值开始正偏移的位置(图10)[5,10]。因此,结合东秋剖面生物在ME中变化情况以及已有的东、西特提斯的观测结果,本研究支持前人观点,即ME期间有机物的持续埋藏可能逐渐降低了大气中的CO2浓度,从而改善SPBE期间由于大量的CO2注入可能引发的全球变暖、海洋酸化以及缺氧等不利于浅水底栖生物生存的环境条件,逐步促进更稳定的生态系统的形成和碳酸盐台地的广泛发育[5,10]。总的来看,ME期间Lithiotis Fauna、有孔虫、腹足、藻类、海百合在整个特提斯热带/亚热带保持发展、扩散和繁盛,表明此期间有机碳的埋藏创造了更加富氧以及中—贫营养条件,有利于后生生物的生存和繁盛[5]。另一个因素可能是在SPBE结束时全球海平面上升[17⁃19],这导致了浅水碳酸盐台地环境的扩张,有利于Lithiotis Fauna及其伴生底栖生物的繁殖和扩散。

图 10 普林斯巴晚期碳同位素与Lithiotis Fauna的全球对比
Figure 10. Global correlation of the carbon isotopic record and Lithiotis Fauna of the Late Pliensbachian
-
在MSBE时期,有孔虫种属和数量明显减少,其中共10种大型底栖有孔虫消失(图2,7)。另外,在此期间东秋剖面的生屑颗粒含量整体呈下降趋势,而且Lithiotis Fauna数量和长度整体也呈现下降的趋势。更值得注意的是,东秋剖面碳同位素负偏移的最低值段(21.5~23.5 m)和灭绝阶段生屑含量最小值之间存在明显同步性(图7)。这些证据表明,碳同位素扰动事件与生物危机之间存在明显联系,但是具体的因素仍需要进行更多的研究。同样地,特提斯喜马拉雅中段卧龙剖面在MSBE开始时部分特征有孔虫灭绝以及生屑颗粒的含量呈下降趋势,并达到最小值[10]。这一事件在时间上与特提斯西部和泛大洋(Panthalassa)东北部margaritatus菊石带中的gibbosus菊石亚带生物危机大致一致[67,88]。另外,Trento碳酸盐台地(意大利)Viote剖面碳同位素负偏移的最低值区域(72~81 m)与Lithiotis Fauna的出现频率最低区域存在明显的同步性(图10c)。这些关联可能表明MSBE时期存在全球生物危机。多个研究表明全球海平面下降会摧毁部分生物的栖息地,海水盐度增加,进而导致海洋生产力下降,从而使得生物多样性和数量的下降[28,89]。正如4.2所述,MSBE期间存在海平面下降,特别是特提斯喜马拉雅Kioto碳酸盐台地(图10b)[10,20],可能导致此次生物危机。然而关于该事件的精确记录,尤其在碳酸盐台地上的记录仍然极少,具体的生物环境响应和成因机制仍有待深入探索。在MSBE之后,台地生物(特别是LBF和Lithiotis Fauna)数量并未恢复到之前的状态,可能是这种环境变化仍未完全恢复。Lithiotis Fauna在西特提斯(意大利和摩洛哥)和东特提斯(西藏)的普林斯巴期浅水碳酸盐台地的分布情况,虽然存在大小、出现的频率以及种类的改变,但直到T-OAE引起的相关的环境和气候变化才完全破坏其生存条件[52]。
-
(1) 提供了特提斯喜马拉雅西段Kioto浅水碳酸盐台地东秋剖面早侏罗世普林斯巴晚期—托阿尔(最早)期高分辨率有孔虫生物地层、碳同位素地层、微相特征以及生屑变化等数据。共识别出3个有孔虫带:Bosniella oenensis⁃Cyclorbitosella tibetica(?)带,Streptocyclammina liasica带和Siphoval vulina sp.A带。因此,东秋剖面地层主体属于普林斯巴阶上部,顶部延伸至托阿尔阶最底部。
(2) 东秋剖面共划分出9种碳酸盐岩微相,主要为泥晶灰岩、白云石化似球粒粒泥灰岩、生屑似球粒泥粒灰岩、生屑泥粒/粒泥灰岩、内碎屑泥粒—颗粒灰岩、团块颗粒灰岩、Lithiotis砾屑灰岩、生屑颗粒灰岩和鲕粒颗粒灰岩。其特征和组合表明普林斯巴阶上部普普嘎组为浅水碳酸盐台地。时间序列上的微相组合变化表明,东秋剖面是由底部到中部的开阔台地和台地边缘环境逐渐过渡到中上部的局限台地,后又演变到顶部的开阔台地和台地边缘的沉积序列。
(3) 在生物地层基础上,根据碳同位素对比在东秋剖面识别出以碳同位素正偏移为特征的margaritatus菊石带事件(ME)和以碳同位素负偏移为特征的margaritatus⁃spinatum界线事件(MSBE)两次事件,进一步证明这两次碳同位素扰动具有全球性特征。在ME期间,生屑颗粒含量呈现增加的趋势、大型底栖有孔虫的种属和数量保持较高的水平,尤其是造礁双壳Lithiotis Fauna的出现并扩散,这与特提斯喜马拉雅中段卧龙剖面和西特提斯观察到的情况类似。这可能是因为该时期的有机质的大量埋藏消耗了SPBE期间产生的大量CO2,从而创造了更适合后生生物生存的海洋环境条件。在MSBE期间,生物丰度(Lithiotis Fauna)和多样性降低以及部分大型底栖有孔虫灭绝,表明该时期存在着生物危机,可能与海平面下降有关。在MSBE之后,生物特别是大型底栖有孔虫和Lithiotis Fauna并未呈现ME时期的状态,可能是MSBE引起这种环境变化仍未恢复。总的来说,普林斯巴晚期碳循环扰动和台地的发展和演化研究仍然严重不足,仍需要未来更多的工作去探索。
Sedimentary and Biotic Response of Kioto Carbonate Platform to the Late Pliensbachian (Early Jurassic) Carbon-Isotope Perturbations in Southern Tibet
-
摘要: 目的 早侏罗世发生了多次重要的碳循环扰动,并伴随气候和环境的重大变化。然而,现有的研究主要集中于西特提斯和欧洲北部,对东特提斯碳循环扰动事件以及定量探究事件对生物影响的相关报道较少。 方法 利用东特提斯藏南Kioto碳酸盐台地普林斯巴晚期—托阿尔(最早)期的生物地层学(大型底栖有孔虫)、沉积学和碳同位素(δ13C)地层学,探讨碳循环扰动以及相应的气候环境变化对该地区的碳酸盐台地生物演化过程影响。 结果 普林斯巴晚期—托阿尔(最早)期大型底栖有孔虫生物地层显示有3个带:Bosniella oenensis-Cyclorbitosella tibetica(?)、Streptocyclammina liasica和Siphovalvulina sp.A。在生物地层基础上,我们识别出以δ13C正偏移为特征的普林斯巴晚期margaritatus菊石带事件(margaritatus zone event,ME)和以δ13C负偏移为特征的margaritatus-spinatum界线事件(margaritatus-spinatum boundary event,MSBE)。 结论 在ME期间,生屑颗粒含量呈增长趋势、大型底栖有孔虫的种属和数量保持较高的水平以及Lithiotis Fauna出现并扩散,这可能表明有机质的大量埋藏消耗了辛涅缪尔晚期—普林斯巴早期产生的大量CO2,从而创造了更适合生物生存的海洋环境条件。在MSBE期间,生屑颗粒含量以及Lithiotis Fauna丰度和长度整体呈下降趋势,并且部分大型底栖有孔虫的灭绝,表明该时期存在着生物危机,可能与海平面下降有关。虽然Lithiotis Fauna存在大小和出现的频率变化,但不可否认的是,Lithiotis Fauna在普林斯巴期整个特提斯浅水碳酸盐台地蓬勃发展,直到T-OAE时期才完全破坏其生存条件。Abstract: Objective During the Early Jurassic, multiple significant perturbations of the carbon-cycle occurred that coincided with abrupt and extreme changes in climate and environment. However, existing research has primarily focused on the western Tethys and northern Europe, with limited attention given to carbon cycle disturbances in the eastern Tethys and the quantitative assessment of their impact on biota. Methods This study examined the biological (larger benthic foraminifera), sedimentological, and carbon-isotope (δ13C) stratigraphy from the southern Tibetan Kioto Platform formed in the southeastern Tethys during the Late Pliensbachian⁃Earliest Toarcian interval to investigate the influence of carbon-cycle perturbations and corresponding climatic and environmental variations on the evolution of carbonate-platform biota in the region. In addition, this study quantitatively analyzed the content of carbonate skeletal grains, the number and species changes of lager benthic foraminifera, and the distribution of Lithiotis Fauna to explore the impact of these events on biota. Results Three foraminiferal zones were recognized: Pliensbachian Bosniella oenensis⁃Cyclorbitosella tibetica (?) and Streptocyclammina liasica, as well as the Earliest Toarcian Siphovalvulina sp. A. According to the sedimentary structure and the quantitative analysis of grains composition and content, this study identified nine carbonate microfacies (MF) from the Pupuga Formation in the Dongqiu section. These microfacies included mudstone (MF1), dolomitic peloidal wackestone (MF2), bioclastic peloidal packstone (MF3), bioclastic wackestone/packstone (MF4), intraclastic packstone⁃grainstone (MF5), lump grainstone (MF6), Lithiotis rudstone (MF7), bioclastic grainstone (MF8), and oolitic grainstone (MF9). The sedimentary microfacies and microfacies assemblages reveal that the Pupuga Formation in the Dongqiu section represented the shallow⁃water carbonate platform deposit, characterized by minimal influence from terrigenous input. Furthermore, temporal variation in microfacies demonstrated that the Dongqiu section provided a comprehensive record of a sedimentary succession, delineating a gradual transition from the open platform and platform margin to the inner platform, succeeded by a swift reversion towards the open platform and platform margin. Based on biostratigraphy, two carbon isotope excursion events in the Late Pliensbachian were identified: the margaritatus zone event (ME) characterized by positive carbon isotope excursion and the margaritatus⁃spinatum zone boundary event (MSBE) with negative carbon isotope excursion as a feature. Conclusions During the ME period, there was an increase in skeletal grain content, maintenance of high species diversity and abundance of larger benthic foraminifera, and the occurrence and spread of Lithiotis Fauna. This may indicate that the persistent burial of organic matter consumed atmospheric pCO2 generated during the Late Sinemurian to Early Pliensbachian interval, creating more suitable marine environmental conditions for biotic survival. In contrast, during the MSBE period, there was a decrease in the abundance and size of Lithiotis Fauna and skeletal grain content, along with the extinction of several index larger benthic foraminifera. This evidence indicates a possible relationship between biotic crises and sea-level fall. Although there were changes in the size and frequency of occurrence of the Lithiotis Fauna, it is undeniable that they flourished throughout the Tethyan shallow carbonate platform during the Pliensbachian interval, and the conditions for skeletal production of carbonates were not completely destroyed until the Toarcian oceanic anoxic event.
-
图 1 特提斯喜马拉雅构造背景和早侏罗世研究区地质背景
Figure 1. Tethys Himalayan tectonic setting and the Early Jurassic geological setting of the study area
图 2 特提斯喜马拉雅西段札达县东秋剖面岩性柱,有孔虫分布和生物地层学框架
Figure 2. Lithological log, foraminiferal distribution, and biostratigraphical framework in the Dongqiu section of Zanda county, western segment of the tethys Himalaya
图 3 札达县东秋剖面普林斯巴晚期—托阿尔(最早)期有孔虫组合图
Figure 3. Late Pliensbachian⁃Earliest Toarcian foraminiferal assemblage in the Dongqiu section of Zanda county
图 4 札达县东秋剖面普普嘎组地层和微相分布
Figure 4. Stratigraphy and microfacies distribution in the Pupuga Formation of the Dongqiu section, Zanda county
图 5 札达县东秋剖面典型微相类型和野外Lithiotis Fauna变化
Figure 5. Typical microfacies types and Lithiotis Fauna changes in the Dongqiu section of Zanda county
图 6 札达县东秋剖面δ13Corg、全岩δ13Ccarb和δ18Ocarb
Figure 6. δ13Corg, bulk δ13Ccarb, and δ18Ocarb in the Dongqiu section of Zanda county
图 7 札达县东秋剖面生屑变化与碳同位素曲线的关系
Figure 7. Relationship between bioclastic change and carbon isotope curve in the Dongqiu section of Zanda county
图 8 (a)δ13Ccarb⁃δ13Corg相关性图解;(b)δ13Ccarb⁃δ18Ocarb相关性图解
Figure 8. (a) Correlation between δ13Ccarb and δ13Corg; (b) correlation between δ13Ccarb and δ18Ocarb
图 9 普林斯巴晚期碳同位素记录的全球对比
Figure 9. Global correlation of the carbon isotopic record of the Late Pliensbachian
-
[1] Jenkyns H C. Geochemistry of oceanic anoxic events[J]. Geochemistry, Geophysics, Geosystems, 2010, 11(3): Q03004. [2] Foster G L, Hull P, Lunt D J, et al. Placing our current ‘hyperthermal’ in the context of rapid climate change in our geological past[J]. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 2018, 376(2130): 20170086. [3] Hu F Z, Fu X G, Lin L, et al. Marine Late Triassic-Jurassic carbon-isotope excursion and biological extinction records: New evidence from the Qiangtang Basin, eastern tethys[J]. Global and Planetary Change, 2020, 185: 103093. [4] Korte C, Hesselbo S P. Shallow marine carbon and oxygen isotope and elemental records indicate icehouse-greenhouse cycles during the Early Jurassic[J]. Paleoceanography, 2011, 26(4): PA4219. [5] Franceschi M, Dal Corso J, Posenato R, et al. Early Pliensbachian (Early Jurassic) C-isotope perturbation and the diffusion of the Lithiotis Fauna: Insights from the western tethys[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 410: 255-263. [6] Franceschi M, Dal Corso J, Cobianchi M, et al. Tethyan carbonate platform transformations during the Early Jurassic (Sinemurian-Pliensbachian, southern Alps): Comparison with the Late Triassic Carnian Pluvial Episode[J]. GSA Bulletin, 2019, 131(7/8): 1255-1275. [7] Baghli H, Mattioli E, Spangenberg J E, et al. Early Jurassic climatic trends in the south-tethyan margin[J]. Gondwana Research, 2020, 77: 67-81. [8] Storm M S, Hesselbo S P, Jenkyns H C, et al. Orbital pacing and secular evolution of the Early Jurassic carbon cycle[J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(8): 3974-3982. [9] Schöllhorn I, Adatte T, Charbonnier G, et al. Pliensbachian environmental perturbations and their potential link with volcanic activity: Swiss and British geochemical records[J]. Sedimentary Geology, 2020, 406: 105665. [10] Han Z, Hu X M, Boudagher-Fadel M, et al. Early Jurassic carbon-isotope perturbations in a shallow-water succession from the tethys Himalaya, southern hemisphere[J]. Newsletters on Stratigraphy, 2021, 54(4): 461-481. [11] Littler K, Hesselbo S P, Jenkyns H C. A carbon-isotope perturbation at the Pliensbachian-Toarcian boundary: Evidence from the Lias Group, NE England[J]. Geological Magazine, 2010, 147(2): 181-192. [12] Peti L, Thibault N, Clémence M E, et al. Sinemurian-Pliensbachian calcareous nannofossil biostratigraphy and organic carbon isotope stratigraphy in the Paris Basin: Calibration to the ammonite biozonation of NW Europe[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 468: 142-161. [13] Mercuzot M, Pellenard P, Durlet C, et al. Carbon-isotope events during the Pliensbachian (Lower Jurassic) on the African and European margins of the NW Tethyan Realm[J]. Newsletters on Stratigraphy, 2020, 53(1): 41-69. [14] Suan G, Mattioli E, Pittet B, et al. Secular environmental precursors to Early Toarcian (Jurassic) extreme climate changes[J]. Earth and Planetary Science Letters, 2010, 290(3/4): 448-458. [15] Korte C, Hesselbo S P, Ullmann C V, et al. Jurassic climate mode governed by ocean gateway[J]. Nature Communications, 2015, 6(1): 10015. [16] Ruhl M, Hesselbo S P, Hinnov L, et al. Astronomical constraints on the duration of the Early Jurassic Pliensbachian stage and global climatic fluctuations[J]. Earth and Planetary Science Letters, 2016, 455: 149-165. [17] Legarreta L, Uliana M A. The Jurassic succession in west-central Argentina: Stratal patterns, sequences and paleogeographic evolution[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1996, 120(3/4): 303-330. [18] Hesselbo S P, Jenkyns H C. British Lower Jurassic sequence stratigraphy[M]//de Graciansky P C, Hardenbol J, Jacquin T, et al. Mesozoic and Cenozoic sequence stratigraphy of European Basins. Tulsa (Oklahoma): SEPM Society for Sedimentary Geology, 1999: 561-581. [19] Haq B U. Jurassic sea-level variations: A reappraisal[J]. GSA Today, 2018, 28(1): 4-10. [20] Han Z, Hu X M, Li J, et al. Jurassic carbonate microfacies and relative sea-level changes in the tethys Himalaya (southern Tibet)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2016, 456: 1-20. [21] Woodfine R G, Jenkyns H C, Sarti M, et al. The response of two tethyan carbonate platforms to the early Toarcian (Jurassic) oceanic anoxic event: Environmental change and differential subsidence[J]. Sedimentology, 2008, 55(4): 1011-1028. [22] Franceschi M, Jin X, Shi Z Q, et al. High-resolution record of multiple organic carbon-isotope excursions in lacustrine deposits of upper Sinemurian through Pliensbachian (Early Jurassic) from the Sichuan Basin, China[J]. GSA Bulletin, 2023, 135(1/2): 3-17. [23] Krencker F N, Fantasia A, Danisch J, et al. Two-phased collapse of the shallow-water carbonate factory during the Late Pliensbachian-Toarcian driven by changing climate and enhanced continental weathering in the northwestern Gondwana margin[J]. Earth-Science Reviews, 2020, 208: 103254. [24] Fraser N M, Bottjer D J, Fischer A G. Dissecting “Lithiotis” bivalves: Implications for the Early Jurassic reef eclipse[J]. Palaios, 2004, 19(1): 51-67. [25] Posenato R, Masetti D. Environmental control and dynamics of Lower Jurassic bivalve build-ups in the Trento Platform (southern Alps, Italy)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2012, 361-362: 1-13. [26] Leinfelder R R, Schmid D U, Nose M, et al. Jurassic reef patterns-The expression of a changing globe[M]//Kiessling W, Flügel E, Golonka J. Phanerozoic Reef Patterns. Tulsa (Oklahoma): SEPM Society for Sedimentary Geology, 2002: 465-520. [27] Trecalli A, Spangenberg J, Adatte T, et al. Carbonate platform evidence of ocean acidification at the onset of the early Toarcian oceanic anoxic event[J]. Earth and Planetary Science Letters, 2012, 357-358(4): 214-225. [28] Posenato R, Bassi D, Trecalli A, et al. Taphonomy and evolution of Lower Jurassic lithiotid bivalve accumulations in the Apennine Carbonate Platform (southern Italy)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2018, 489: 261-271. [29] 尹安. 喜马拉雅造山带新生代构造演化:沿走向变化的构造几何形态、剥露历史和前陆沉积的约束[J]. 地学前缘,2006,13(5):416-515. Yin An. Cenozoic tectonic evolution of the Himalayan orogen as constrained by along-strike variation of structural geometry, exhumation history, and forel and sedimentation[J]. Earth Science Frontiers, 2006, 13(5): 416-515. [30] Yin A, Harrison T M. Geologic evolution of the Himalayan-Tibetan orogen[J]. Annual Review of Earth and Planetary Sciences, 2000, 28(1): 211-280. [31] Burg J P, Bouilhol P. Timeline of the South Tibet-Himalayan belt: The geochronological record of subduction, collision, and underthrusting from zircon and monazite U-Pb ages[J]. Canadian Journal of Earth Sciences, 2019, 56(12): 1318-1332. [32] Golonka J. Phanerozoic paleoenvironment and paleolithofacies maps. Mesozoic[J]. Geologia, 2007, 33: 211-264 [33] Han Z, Hu X M, Kemp D B, et al. Carbonate-platform response to the Toarcian Oceanic Anoxic Event in the southern hemisphere: Implications for climatic change and biotic platform demise[J]. Earth and Planetary Science Letters, 2018, 489: 59-71. [34] Jadoul F, Berra F, Garzanti E. The tethys Himalayan passive margin from Late Triassic to Early Cretaceous (south Tibet)[J]. Journal of Asian Earth Sciences, 1998, 16(2/3): 173-194. [35] Garzanti E. Stratigraphy and sedimentary history of the Nepal Tethys Himalaya passive margin[J]. Journal of Asian Earth Sciences, 1999, 17(5/6): 805-827. [36] 阴家润,万晓樵. 侏罗纪菊石形态—特提斯喜马拉雅海的深度标志[J]. 古生物学报,1996,35(6):734-751. Yin Jiarun, Wan Xiaoqiao. Jurassic ammonite morphotypes as water-depth indicator of tethys-Himalaya sea[J]. Acta Palaeontologica Sinica, 1996, 35(6): 734-751. [37] Sciunnach D, Garzanti E. Subsidence history of the Tethys Himalaya[J]. Earth-Science Reviews, 2012, 111(1/2): 179-198. [38] 李金和,孙苑迪,夏祥. 西藏札达地区早侏罗世双壳类Lithiotis生物岩礁的发现及其地质意义[J]. 地球,2013(8):52-53. Li Jinhe, Sun Yuandi, Xia Xiang. Discovery of the Early Jurassic Lithiotis (Bivalvia) bioherm in Zhada area in Tibet and its geological implications[J]. The Earth, 2013(8): 52-53. [39] 张双增,季金和,张宏宝,等. 中华人民共和国区域地质调查报告:日新幅(H44C001001)、札达县幅(H44C002001)、姜叶马幅(H44C002002)比例尺1∶250000 [M]. 北京:地质出版社,2015:1-224. Zhang Shuangzeng, Ji Jinhe, Zhang Hongbao, et al. Regional geological survey report of the People's Republic of China: Rixin sheet (H44C001001), Zanda county sheet (H44C002001), Jiangyema sheet (H44C002002), scale 1:250,000 [M]. Beijing: Geological Publishing House, 2015: 1-224. [40] Wignall P B, Hallam A, Newton R J, et al. An eastern tethyan (Tibetan) record of the Early Jurassic (Toarcian) mass extinction event[J]. Geobiology, 2006, 4(3): 179-190. [41] Newton R J, Reeves E P, Kafousia N, et al. Low marine sulfate concentrations and the isolation of the European epicontinental sea during the Early Jurassic[J]. Geology, 2011, 39(1): 7-10.. [42] Loeblich A R, Tappan H. Foraminiferal genera and their classification[M]. New York: Springer, 1988. [43] Boudagher-Fadel M K. Evolution and geological significance of larger benthic foraminifera[M]. Amsterdam: Elsevier, 2008: 1-544. [44] Boudagher-Fadel M K. Evolution and geological significance of larger benthic foraminifera[M]. London: UCL Press, 2018: 1-693. [45] Riding R. Structure and composition of organic reefs and carbonate mud mounds: Concepts and categories[J]. Earth-Science Reviews, 2002, 58(1/2): 163-231. [46] Trygonis V, Sini M. PhotoQuad: A dedicated seabed image processing software, and a comparative error analysis of four photoquadrat methods[J]. Journal of Experimental Marine Biology and Ecology, 2012, 424-425: 99-108. [47] Solomon M. Counting and sampling errors in modal analysis by point counter[J]. Journal of Petrology, 1963, 4(3): 367-382. [48] Eriksson M E, Lindskog A, Calner M, et al. Biotic dynamics and carbonate microfacies of the conspicuous Darriwilian (Middle Ordovician) 'Täljsten' interval, south-central Sweden[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2012, 367-368: 89-103. [49] Flügel E. Microfacies of carbonate rocks: Analysis, interpretation and application[M]. 2nd ed. Berlin Heidelberg: Springer, 2010: 1-984. [50] Barattolo F, Romano R. Shallow carbonate platform bioevents during the Upper Triassic-Lower Jurassic: An evolutive interpretation[J]. Italian Journal of Geosciences, 2005, 124(1): 123-142. [51] Boudagher-Fadel M K, Bosence D W J. Early Jurassic benthic foraminiferal diversification and biozones in shallow-marine carbonates of western tethys[J]. Senckenbergiana lethaea, 2007, 87(1): 1-39. [52] Brame H M R, Martindale R C, Ettinger N P, et al. Stratigraphic distribution and paleoecological significance of Early Jurassic (Pliensbachian-Toarcian) lithiotid-coral reefal deposits from the central High Atlas of Morocco[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2019, 514: 813-837. [53] Oehlert A M, Swart P K. Interpreting carbonate and organic carbon isotope covariance in the sedimentary record[J]. Nature Communications, 2014, 5: 4672. [54] Suan G, van de Schootbrugge B, Adatte T, et al. Calibrating the magnitude of the Toarcian carbon cycle perturbation[J]. Paleoceanography, 2015, 30(5): 495-509. [55] Fike D A, Grotzinger J P, Pratt L M, et al. Oxidation of the Ediacaran ocean[J]. Nature, 2006, 444(7120): 744-747 [56] Grotzinger J P, Fike D A, Fischer W W. Enigmatic origin of the largest-known carbon isotope excursion in Earth's history[J]. Nature Geoscience, 2011, 4(5): 285-292. [57] Rose C V, Swanson-Hysell N L, Husson J M, et al. Constraints on the origin and relative timing of the Trezona δ13C anomaly below the end-Cryogenian glaciation[J]. Earth and Planetary Science Letters, 2012, 319-320: 241-250. [58] Brasier M D, Magaritz M, Corfield R, et al. The carbon-and oxygen-isotope record of the Precambrian–Cambrian boundary interval in China and Iran and their correlation[J]. Geological Magazine, 1990, 127(4): 319-332. [59] Kaufman A J, Knoll A H. Neoproterozoic variations in the C-isotopic composition of seawater: Stratigraphic and biogeochemical implications[J]. Precambrian Research, 1995, 73(1/2/3/4): 27-49. [60] Knoll A H, Hayes J M, Kaufman A J, et al. Secular variation in carbon isotope ratios from Upper Proterozoic successions of Svalbard and East Greenland[J]. Nature, 1986, 321(6073): 832-838. [61] Laporte D F, Holmden C, Patterson W P, et al. Local and global perspectives on carbon and nitrogen cycling during the Hirnantian glaciation[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2009, 276(1/2/3/4): 182-195. [62] Swanson-Hysell N L, Rose C V, Calmet C C, et al. Cryogenian glaciation and the onset of carbon-isotope decoupling[J]. Science, 2010, 328(5978): 608-611. [63] Meyer K M, Yu M, Lehrmann D, et al. Constraints on Early Triassic carbon cycle dynamics from paired organic and inorganic carbon isotope records[J]. Earth and Planetary Science Letters, 2013, 361: 429-435. [64] van de Schootbrugge B, Mcarthur J M, Bailey T R, et al. Toarcian oceanic anoxic event: An assessment of global causes using belemnite C isotope records[J]. Paleoceanography, 2005, 20(3): PA3008. [65] Rosales I, Quesada S, Robles S. Geochemical arguments for identifying second-order sea-level changes in hemipelagic carbonate ramp deposits[J]. Terra Nova, 2006, 18(4): 233-240. [66] Price G D, Baker S J, Vandevelde J, et al. High-resolution carbon cycle and seawater temperature evolution during the Early Jurassic (Sinemurian-Early Pliensbachian)[J]. Geochemistry, Geophysics, Geosystems, 2016, 17(10): 3917-3928. [67] Caruthers A H, Smith P L, Gröcke D R. The Pliensbachian-Toarcian (Early Jurassic) extinction: A North American perspective[M]//Keller G, Kerr A C. Volcanism, impacts, and mass extinctions: Causes and effects. Boulder: Geological Society of America, 2014, 505: 225-243. [68] de Lena L F, Taylor D, Guex J, et al. The driving mechanisms of the carbon cycle perturbations in the Late Pliensbachian (Early Jurassic)[J]. Scientific Reports, 2019, 9(1): 18430. [69] Silva R L, Duarte L V, Comas-Rengifo M J, et al. Update of the carbon and oxygen isotopic records of the Early-Late Pliensbachian (Early Jurassic, ~187Ma): Insights from the organic-rich hemipelagic series of the Lusitanian Basin (Portugal)[J]. Chemical Geology, 2011, 283(3/4): 177-184. [70] Silva R L, Duarte L V. Organic matter production and preservation in the Lusitanian Basin (Portugal) and Pliensbachian climatic hot snaps[J]. Global and Planetary Change, 2015, 131: 24-34. [71] Han Z, Hu X M, He T C, et al. Early Jurassic long-term oceanic sulfur-cycle perturbations in the Tibetan Himalaya[J]. Earth and Planetary Science Letters, 2022, 578: 117261. [72] Harazim D, van de Schootbrugge B, Sorichter K, et al. Spatial variability of watermass conditions within the European Epicontinental Seaway during the Early Jurassic (Pliensbachian-Toarcian)[J]. Sedimentology, 2013, 60(2): 359-390. [73] Gómez J J, Comas-Rengifo M J, Goy A. Palaeoclimatic oscillations in the Pliensbachian (Lower Jurassic) of the Asturian Basin (northern Spain)[J]. Climate of the Past Discussions, 2015, 11(4): 4039-4076. [74] Arabas A, Schlögl J, Meister C. Early Jurassic carbon and oxygen isotope records and seawater temperature variations: Insights from marine carbonate and belemnite rostra (Pieniny Klippen Belt, Carpathians)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 485: 119-135. [75] Bougeault C, Pellenard P, Deconinck J F, et al. Climatic and palaeoceanographic changes during the Pliensbachian (Early Jurassic) inferred from clay mineralogy and stable isotope (C-O) geochemistry (NW Europe)[J]. Global and Planetary Change, 2017, 149: 139-152. [76] Price G D. The evidence and implications of polar ice during the Mesozoic[J]. Earth-Science Reviews, 1999, 48(3): 183-210. [77] Rogov M A, Zakharov V A. Jurassic and Lower Cretaceous glendonite occurrences and their implication for Arctic paleoclimate reconstructions and stratigraphy[J]. Earth Science Frontiers, 2010, 17(Suppl.1): 345-347. [78] Teichert B M A, Luppold F W. Glendonites from an Early Jurassic methane seep:Climate or methane indicators?[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2013, 390: 81-93. [79] Philippe M, Puijalon S, Suan G, et al. The palaeolatitudinal distribution of fossil wood genera as a proxy for European Jurassic terrestrial climate[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 466: 373-381. [80] Steinthorsdottir M, Vajda V. Early Jurassic (Late Pliensbachian) CO2 concentrations based on stomatal analysis of fossil conifer leaves from eastern Australia[J]. Gondwana Research, 2015, 27(3): 932-939. [81] Ruebsam W, Mayer B, Schwark L. Cryosphere carbon dynamics control early Toarcian global warming and sea level evolution[J]. Global and Planetary Change, 2019, 172: 440-453. [82] Hesselbo S P, Jenkyns H C, Duarte L V, et al. Carbon-isotope record of the Early Jurassic (Toarcian) Oceanic Anoxic Event from fossil wood and marine carbonate (Lusitanian Basin, Portugal)[J]. Earth and Planetary Science Letters, 2007, 253(3/4): 455-470. [83] Ait-Itto F Z, Price G D, Addi A A, et al. Bulk-carbonate and belemnite carbon-isotope records across the Pliensbachian-Toarcian boundary on the northern margin of Gondwana (Issouka, Middle Atlas, Morocco)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 466: 128-136. [84] Izumi K, Kemp D B, Itamiya S, et al. Sedimentary evidence for enhanced hydrological cycling in response to rapid carbon release during the early Toarcian oceanic anoxic event[J]. Earth and Planetary Science Letters, 2018, 481: 162-170. [85] Fantasia A, Föllmi K B, Adatte T, et al. The Toarcian oceanic anoxic event in southwestern Gondwana: An example from the Andean Basin, northern Chile[J]. Journal of the Geological Society, 2018, 175(6): 883-902. [86] Bodin S, Fantasia A, Krencker F N, et al. More gaps than record! A new look at the Pliensbachian/Toarcian boundary event guided by coupled chemo-sequence stratigraphy[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2023, 610: 111344. [87] Krencker F N, Fantasia A, El Ouali M, et al. The effects of strong sediment-supply variability on the sequence stratigraphic architecture: Insights from early Toarcian carbonate factory collapses[J]. Marine and Petroleum Geology, 2022, 136: 105469. [88] Dera G, Neige P, Dommergues J L, et al. High-resolution dynamics of Early Jurassic marine extinctions: The case of Pliensbachian-Toarcian ammonites (Cephalopoda)[J]. Journal of the Geological Society, 2010, 167(1): 21-33. [89] Takahashi A. Diversity changes in Cretaceous inoceramid bivalves of Japan[J]. Paleontological Research, 2005, 9(3): 217-232. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 968
- HTML全文浏览量: 235
- PDF下载量: 99
- 被引次数: 0

